七大鐵死亡檢測(cè)方法及其典型實(shí)驗(yàn)案例拆解
Section.02
鐵死亡檢測(cè)方法
檢測(cè)鐵死亡的方法包括檢測(cè)代謝反應(yīng) (如脂代謝,鐵代謝和 GSH 相關(guān)代謝) 相關(guān)標(biāo)志物含量變化,細(xì)胞形態(tài)變化和分子蛋白含量/活性變化。
1. 過(guò)氧化脂質(zhì)檢測(cè)
脂質(zhì)過(guò)氧化是鐵死亡的核心驅(qū)動(dòng)因素,過(guò)度的脂質(zhì)過(guò)氧化通過(guò)產(chǎn)生有毒的磷脂氫過(guò)氧化物 (PLOOH),在促進(jìn)鐵死亡中起著重要作用,因此過(guò)氧化脂質(zhì)可用于細(xì)胞鐵死亡的檢測(cè)。
在對(duì)骨關(guān)節(jié)炎中軟骨細(xì)胞鐵死亡的研究中,為了證實(shí) SM@ApoFn nanocage 具有抑制細(xì)胞鐵死亡的功能,作者用脂溶性比率型熒光探針 BODIPY 581/591 C11 來(lái)評(píng)估細(xì)胞內(nèi)脂質(zhì)過(guò)氧化水平。如圖所示,BODIPY 581/591 C11 (10 μM) 探針?lè)治?IL-1β 預(yù)處理 12 小時(shí)后,各組給予 SM04690、ApoFn 和 SM@ApoFn 處理 24 h 的軟骨細(xì)胞中細(xì)胞膜脂質(zhì)的氧化和還原狀態(tài),其中 SM@ApoFn 減輕 IL-1β 引起的脂質(zhì)過(guò)氧化。
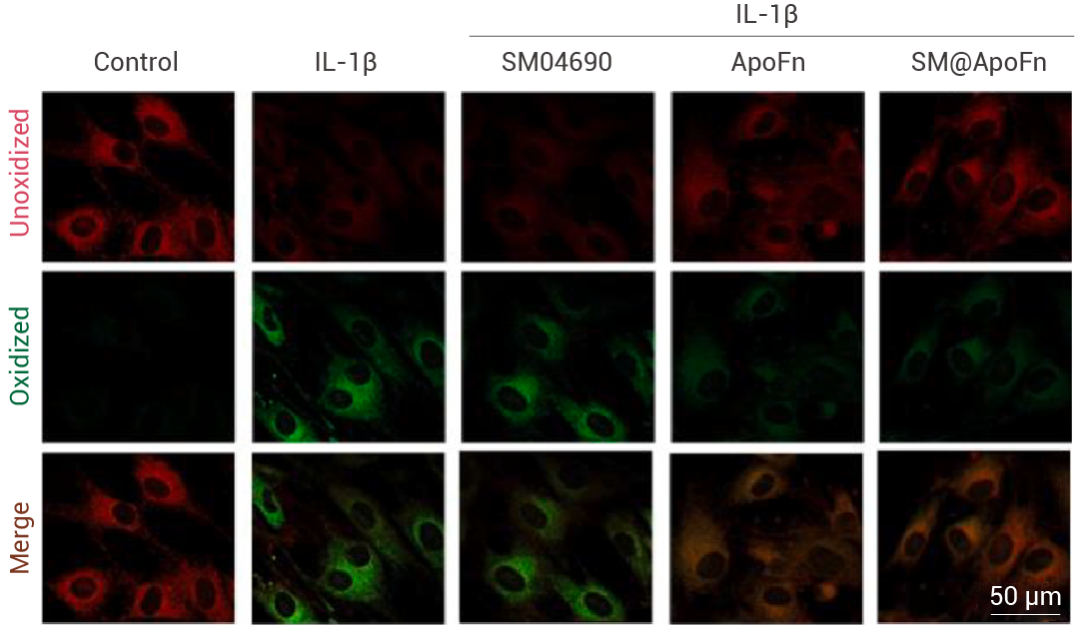 圖 1. SM@ApoFn 減輕 IL-1β 引起的脂質(zhì)過(guò)氧化[1]。
圖 1. SM@ApoFn 減輕 IL-1β 引起的脂質(zhì)過(guò)氧化[1]。 BODIPY 581/591 C11 (10 μM) 探針?lè)治?IL-1β 預(yù)處理 12 小時(shí)后,各組給予 SM04690/ApoFn 和 SM@ApoFn 處理 24 h 的軟骨細(xì)胞中細(xì)胞膜脂質(zhì)的氧化和還原狀態(tài)。
2. 活性氧檢測(cè)
活性氧 (ROS) 在鐵死亡過(guò)程中不僅促進(jìn)脂質(zhì)過(guò)氧化和細(xì)胞損傷,還抑制抗氧化信號(hào)通路,是鐵死亡 檢 測(cè) 的 重 要 生 物 標(biāo) 志 物。
在研究不同谷胱甘肽過(guò)氧化物酶 4 (GPX4) 抑制劑的納米藥物開(kāi)發(fā)中,用 DCFH-DA 熒光探針檢測(cè)不同納米藥物處理的 HT-1080 細(xì)胞中總的 ROS 水平。其中納米藥物 2b 對(duì) GPX4 有較好的抑制效果,導(dǎo)致細(xì)胞內(nèi)的 ROS 水平顯著升高。
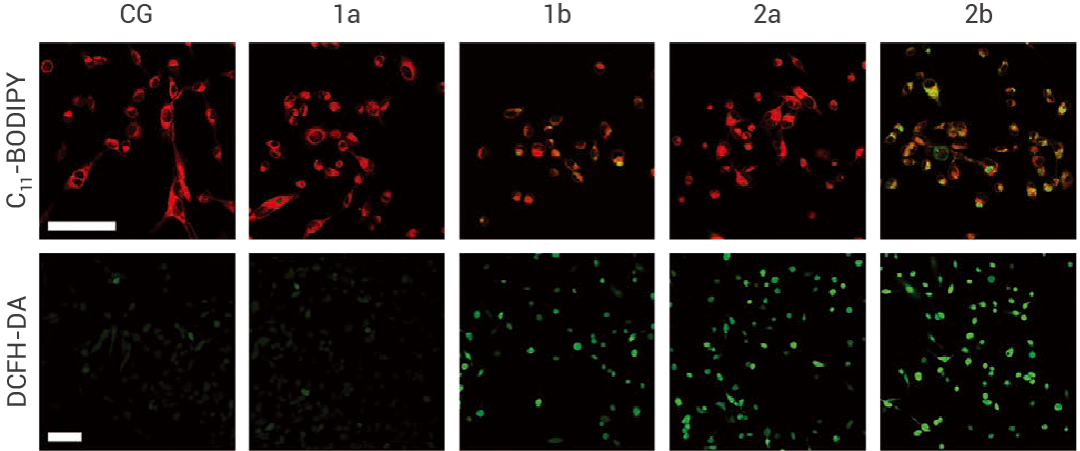 圖 2. 不同納米藥物處理的 HT-1080 細(xì)胞中的總 ROS 水平檢測(cè)[2]。
圖 2. 不同納米藥物處理的 HT-1080 細(xì)胞中的總 ROS 水平檢測(cè)[2]。
用 C11-BODIPY 和 DCFH-DA 熒光探針檢測(cè)不同納米藥物處理的 HT-1080 細(xì)胞內(nèi)脂質(zhì)過(guò)氧化和總 ROS。
3. 過(guò)氧化脂質(zhì)衍生物檢測(cè)
脂質(zhì)過(guò)氧化反應(yīng)過(guò)程中,大量的脂質(zhì)過(guò)氧化初級(jí)產(chǎn)物在氧化反應(yīng)中逐漸分解為一系列復(fù)雜的醛類化合物,包括 MDA,4-HNE,丙醛和己醛等。這些活性醛類物質(zhì)能夠攻擊蛋白質(zhì)等生物大分子,形成加合物,從而引發(fā)細(xì)胞的進(jìn)一步損傷。
在研究真核翻譯起始因子 EIF4E 通過(guò)控制脂質(zhì)過(guò)氧化作為鐵死亡敏感性的決定因素的研究中。科研人員為了研究 4HNE 產(chǎn)量增加是否會(huì)影響反饋回路,從而降低細(xì)胞對(duì)鐵死亡的敏感性。結(jié)果表明,用 RSL3 處理后,ALDH1B1 的過(guò)表達(dá)限制了 WT 和 EIF4E 過(guò)表達(dá)的 Calu-1 細(xì)胞中 4HNE 的積累,這表明 ALDH1B1 在限制鐵死亡期間細(xì)胞中 4HNE 的產(chǎn)生方面仍然發(fā)揮著重要作用。
需要注意的是,MDA 和 4-HNE 在除鐵死亡外的多種氧化 應(yīng)激狀態(tài)下均可產(chǎn)生,在實(shí)際應(yīng)用中,一般與其他檢測(cè)方法進(jìn)行相互驗(yàn)證,以提高結(jié)果的可靠性和準(zhǔn)確性。
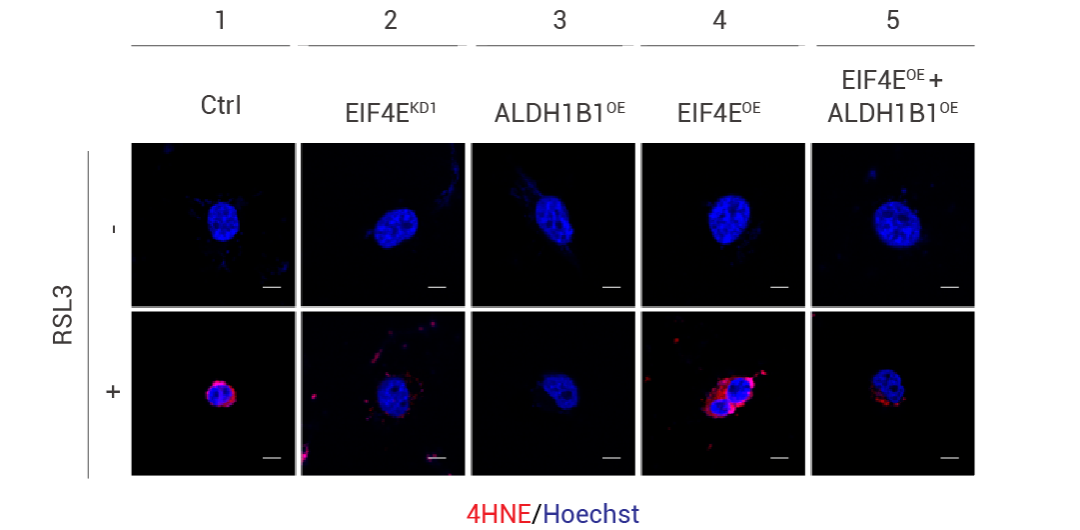 圖 3. 用 RSL3(0.5 μM,4 小時(shí))處理不同過(guò)表達(dá)體系的 Calu-1 細(xì)胞中 4HNE 染色的免疫熒光分析[3]。
圖 3. 用 RSL3(0.5 μM,4 小時(shí))處理不同過(guò)表達(dá)體系的 Calu-1 細(xì)胞中 4HNE 染色的免疫熒光分析[3]。
4. 鐵代謝相關(guān)生物標(biāo)志物檢測(cè)
鐵死亡過(guò)程中,細(xì)胞內(nèi)亞鐵離子的積累會(huì)導(dǎo)致氧化應(yīng)激的增加,促進(jìn)脂質(zhì)過(guò)氧化。因此,細(xì)胞鐵含量或者 Fe2+/Fe3+ 比值的測(cè)定可作為鐵死亡監(jiān)測(cè)的重要指標(biāo)。FerroOrange 是一種特異性 Fe2+ 探針,與 Fe2+ 結(jié)合形成橙色熒光復(fù)合物,可通過(guò)熒光顯微鏡或流式細(xì)胞術(shù)檢測(cè) (Ex/Em: 542/572 nm), 從而定量分析細(xì)胞內(nèi) Fe2+ 的濃度。
研究人員在明確 PGS(一種天然產(chǎn)物 P-L)對(duì)腎臟、智力及神經(jīng)保護(hù)的潛在作用,探索 PGS 的確切靶點(diǎn)及作用機(jī)制。在研究 PGS 是否減輕缺氧 BV2 細(xì)胞脂質(zhì)過(guò)氧化物積累引起的鐵死亡實(shí)驗(yàn)中,使用 FerroOrange 染色,PGS 可減輕 BV2 細(xì)胞中缺氧 (HH) 誘導(dǎo)的鐵死亡。
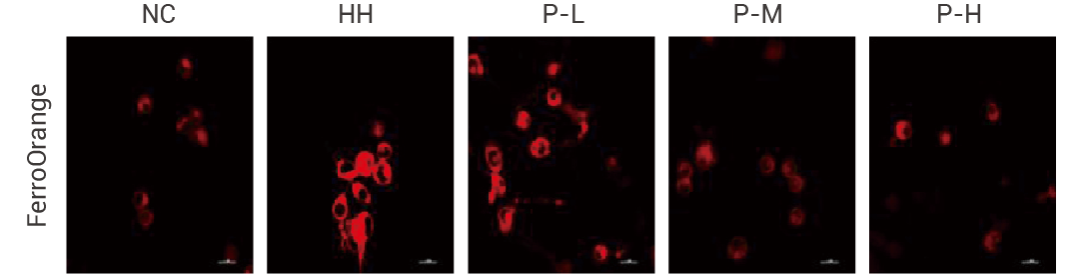 圖 4. PGS (一種天然產(chǎn)物 P-L, 50 μg/mL; P-M, 100 μg/mL; P-H, 200 μg/mL) 可減輕 BV2 細(xì)胞中缺氧 (HH) 誘導(dǎo)的鐵死亡[4]。
圖 4. PGS (一種天然產(chǎn)物 P-L, 50 μg/mL; P-M, 100 μg/mL; P-H, 200 μg/mL) 可減輕 BV2 細(xì)胞中缺氧 (HH) 誘導(dǎo)的鐵死亡[4]。
5. 谷胱甘肽代謝相關(guān)生物標(biāo)志物檢測(cè)
谷胱甘肽 (GSH) 是細(xì)胞內(nèi)主要的抗氧化劑,在鐵死亡過(guò)程中,GSH 水平顯著下降,導(dǎo)致抗氧化能力減弱, 進(jìn)而促進(jìn)脂質(zhì)過(guò)氧化物的積累。因此,檢測(cè) GSH 相關(guān)代謝等指標(biāo)可用于監(jiān)測(cè)鐵死亡的過(guò)程。
鐵死亡在腸缺血再灌注損傷中起著至關(guān)重要的作用,抑制鐵死亡可能為治療該疾病提供新的途徑,研究人員在對(duì)鐵死亡發(fā)生的小鼠腸道缺血再灌注模型中進(jìn)行檢測(cè),再灌注 30 min 后,組織中總鐵和 Fe2+ 含量最高,LPO 產(chǎn)物 MDA 也隨再灌注時(shí)間延長(zhǎng)而積累,還原型 GSH 水平降低,氧化型 GSSG 水平升高。
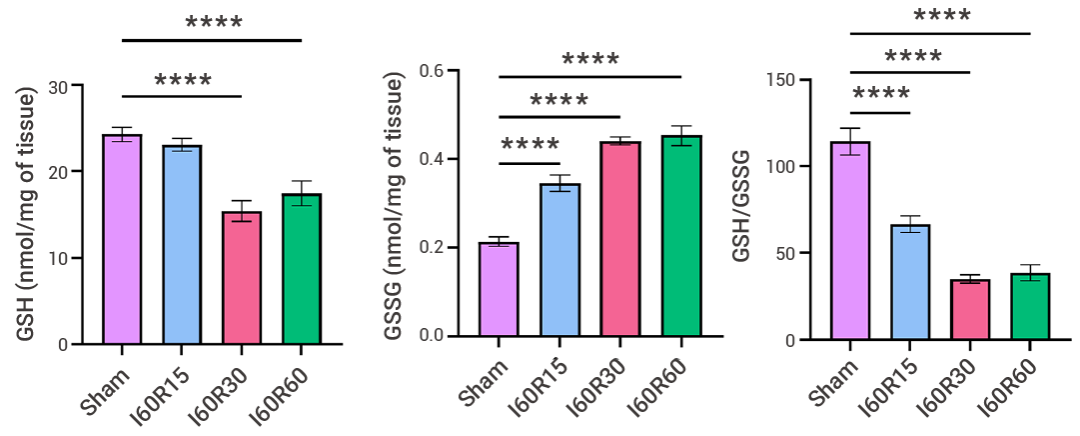 圖 5. 小鼠腸道缺血再灌注模型中,再灌注 30 min 后使用 GSH + GSSG/GSH 檢測(cè)試劑盒評(píng)估腸道組織中谷胱甘肽、GSH、GSSG 和 GSH/GSSG 水平[5]。
圖 5. 小鼠腸道缺血再灌注模型中,再灌注 30 min 后使用 GSH + GSSG/GSH 檢測(cè)試劑盒評(píng)估腸道組織中谷胱甘肽、GSH、GSSG 和 GSH/GSSG 水平[5]。
6. 細(xì)胞和亞細(xì)胞水平的形態(tài)學(xué)檢測(cè)
鐵死亡細(xì)胞的典型形態(tài)學(xué)特征包括質(zhì)膜完整性喪失,細(xì)胞膜破裂,線粒體萎縮,線粒體外膜破裂、嵴減少或缺失、膜密度增加,可通過(guò)透射電子顯微鏡 (TEM) 檢測(cè)。此外,鐵死亡發(fā)生時(shí),也伴隨著線粒體膜通透性增加和線粒體膜電位降低,內(nèi)質(zhì)網(wǎng)粘度增加和內(nèi)質(zhì)網(wǎng)應(yīng)激, 以及溶酶體鐵或一氧化氮 (NO) 的累積。可借助相關(guān)熒光探針,用流式細(xì)胞儀或熒光顯微鏡檢測(cè)。
在研究溴結(jié)構(gòu)域蛋白 4(BRD4)抑制 Erastin 誘導(dǎo)的鐵死亡中,研究人員使測(cè)量了 BRD4 抑制前后細(xì)胞的氧化分解代謝,與 Erastin 處理組相比,當(dāng) BRD4 被其抑制劑 (+)-JQ1 和 PFI-1 抑制時(shí),檢測(cè)到的線粒體膜電位低得多。
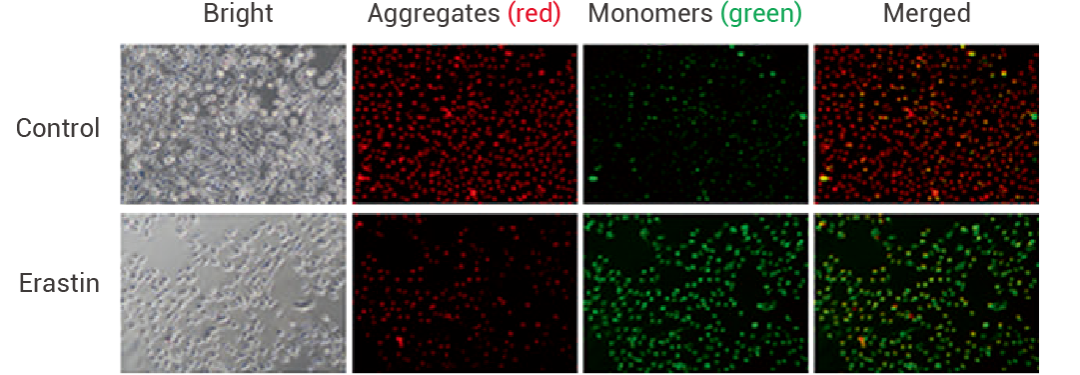 圖 6. Erastin (10 μM, 24 h) 誘導(dǎo) HT1080 鐵死亡,JC-1 檢測(cè)線粒體膜電位變化[6]。
圖 6. Erastin (10 μM, 24 h) 誘導(dǎo) HT1080 鐵死亡,JC-1 檢測(cè)線粒體膜電位變化[6]。
7. 調(diào)控鐵死亡的基因和蛋白的檢測(cè)
鐵死亡主要由鐵依賴性脂質(zhì)過(guò)氧化引起,其調(diào)控機(jī)制包括多種,主要分為過(guò)氧化代謝部分 (如鐵代謝和脂代謝途徑) 和抗氧化部分 (如 System xc--GSH-GPX4 途徑,AIFM2-CoQ10/GCH1-BH4/ ESCRT-III 和 DHODH 途徑)。鐵死亡發(fā)生時(shí),一些關(guān)鍵基因和蛋白表達(dá)水平發(fā)生變化,因此,采用 Western blot / 免疫熒光 / 免疫組織化學(xué) / 實(shí)時(shí)熒光定量 PCR 技術(shù)等方法來(lái)表征鐵死亡的發(fā)生。
在探討鐵死亡是否參與缺氧缺血性腦損傷及其機(jī)制的研究中,通過(guò)阻斷左頸總動(dòng)脈建立 7 日齡雄性 Sprague-Dawley 新生大鼠 HIBD 模型。HIBD 組大鼠的活性氧水平顯著升高,鐵代謝相關(guān)蛋白轉(zhuǎn)鐵蛋白受體 (TFRC)、鐵蛋白重鏈 (FHC)、鐵蛋白輕鏈 (FLC) 的蛋白水平顯著升高,而 SLC7A11、GSH、GPX4 的蛋白水平降低,這些變化導(dǎo)致細(xì)胞抗氧化能力下降、線粒體損傷,引起大腦皮層神經(jīng)元鐵死亡。
 圖 7. 通過(guò)阻斷左頸總動(dòng)脈建立 7 日齡雄性 Sprague-Dawley 新生大鼠 HIBD 模型后 72 小時(shí)腦組織中鐵死亡相關(guān)蛋白的水平[7]。
圖 7. 通過(guò)阻斷左頸總動(dòng)脈建立 7 日齡雄性 Sprague-Dawley 新生大鼠 HIBD 模型后 72 小時(shí)腦組織中鐵死亡相關(guān)蛋白的水平[7]。
Section.03
小結(jié)
今天,小 M 給大家詳細(xì)介紹了鐵死亡的關(guān)鍵指標(biāo)和對(duì)應(yīng)的檢測(cè)方式,檢測(cè)指標(biāo)也呈現(xiàn)出多樣化的特點(diǎn)。鐵死亡收到多條通路的調(diào)控,在實(shí)驗(yàn)檢測(cè)過(guò)程中,各個(gè)通路相輔相成,互相影響,大家在檢測(cè)鐵死亡時(shí),需要組合出擊,多種檢測(cè)方式共同驗(yàn)證,實(shí)驗(yàn)結(jié)果才會(huì)更有說(shuō)服力哦。
|
產(chǎn)品推薦 |
|
比率型脂質(zhì)過(guò)氧化熒光探針;健康細(xì)胞:紅色熒光 (Ex/Em=581/591 nm);鐵死亡細(xì)胞:綠色熒光 (Ex/Em=500/510 nm) |
|
ROS 探針;健康細(xì)胞: 微弱的綠色熒光;鐵死亡細(xì)胞: 更為明亮的綠色熒光 (Ex/Em=488/525 nm) |
|
Fe2+ 探針;橙色熒光 (Ex/Em=542/572 nm) |
|
SLC7A11 (xCT) Antibody (YA006) (HY-P80935) 兔源,用于人、小鼠背景下 WB、IP 實(shí)驗(yàn) |
|
兔源,用于人、小鼠、大鼠背景下 WB、IHC-P、ICC 實(shí)驗(yàn) |
|
一種鐵死亡 (ferroptosis) 誘導(dǎo)劑。Erastin 的鐵死亡誘導(dǎo)機(jī)制與 ROS 和鐵依賴性信號(hào)傳導(dǎo)有關(guān) |
|
一種有效的、選擇性的 ferroptosis 抑制劑,抑制 Erastin 誘導(dǎo)的 HT-1080 細(xì)胞鐵死亡 (EC50=60 nM) |

[1] Yuxin Deng, et al. A novel apoferritin nanocage with ECM promoting, ferroptosis suppressing and inflammation targeting property for osteoarthritis therapy. Chemical Engineering Journal 493 (2024): 152398.
[2] LeLe Zhou, et al. A ferrocene‐functionalized covalent organic framework for enhancing chemodynamic therapy via redox dyshomeostasis. Small 17.32 (2021): 2101368.
[3] Xin Chen, et al. A noncanonical function of EIF4E limits ALDH1B1 activity and increases susceptibility to ferroptosis. Nature communications 13.1 (2022): 6318.
[4] Xianxie Zhang, et al. Ferroptosis pathways: unveiling the neuroprotective power of cistache deserticola phenylethanoid glycosides. Journal of Ethnopharmacology 333 (2024): 118465.
[5] Xingjie Wang, et al. Resveratrol reduces ROS-induced ferroptosis by activating SIRT3 and compensating the GSH/GPX4 pathway. Molecular Medicine 29.1 (2023): 137.
[6] Minghua Yang, et al. Bromodomain-containing protein 4 (BRD4) as an epigenetic regulator of fatty acid metabolism genes and ferroptosis. Cell Death & Disease 13.10 (2022): 912.
[7] Wei Lin, et al. Ferroptosis is involved in hypoxic-ischemic brain damage in neonatal rats. Neuroscience 487 (2022): 131-142.



- Mitomycin C的作用機(jī)制及在腫瘤細(xì)胞系和動(dòng)物模型中的多場(chǎng)景應(yīng)用
- CFDA-SE的作用機(jī)制及在細(xì)胞增殖和體內(nèi)示蹤中的應(yīng)用
- 抑制劑A83-01的作用機(jī)制及在干細(xì)胞與類器官培養(yǎng)中的應(yīng)用
- 2-Deoxy-D-glucose(2-DG)的作用機(jī)制及在糖酵解抑制研究中的作用
- 鈣離子熒光探針Calbryte 520.AM在動(dòng)態(tài)監(jiān)測(cè)中的核心優(yōu)勢(shì)與應(yīng)用解析
- NRF2和AKT激活或FBP1缺失在肝細(xì)胞衰老和肝癌進(jìn)展中的作用
- 多肽的功能和應(yīng)用及其固相合成技術(shù)
- 鐵死亡抑制劑Ferrostatin-1的作用機(jī)制與應(yīng)用
- MCE六月展會(huì)預(yù)告:人工智能大會(huì)&干細(xì)胞研究會(huì)議
- MCE HOLO系列均相免疫分析試劑盒上新5折嘗鮮價(jià)
- MCE直播預(yù)告:PROTAC研究新進(jìn)展和設(shè)計(jì)新思路
- MCE端午雙重福利:手速PK賽和熱門多肽產(chǎn)品滿贈(zèng)禮品
- MCE中國(guó)初夏促銷活動(dòng):染料產(chǎn)品鉅惠55折狂歡
- MCE與你相約第六屆國(guó)際細(xì)胞與基因治療中國(guó)峰會(huì)
- 西安百螢618年中大促,買熒光染料送高通量專用微孔板
- 西美杰成功參加第二十四屆中國(guó)生物制品大會(huì)