
ТЦЦЖ„©Z-VAD-FMKөДЧчУГҷCАнј°‘ӘУГ°ёАэФ”Ҫв
јҡ°ыөтНцЎўҪ№НцәНүДЛАРФөтНцКЗјҡ°ыЛАНцСРҫҝЦРҺЧӮҖҹбйTөДјҡ·ЦоIУтЈ¬ТІКЗЙъОпҫSіЦғИӯhҫі·Җ¶ЁЎўЯMРРҪMҝ—°lУэәНЗеіэ“pӮыјҡ°ыөДЦШТӘҷCЦЖЎЈZ-VAD-FMKЈЁAbMoleЈ¬M3143Ј©КЗТ»ҝо·әCaspaseЈЁpan-CaspaseЈ©ТЦЦЖ„©Ј¬ДЬүтУРР§Чи”ајҡ°ыөтНцЎўҪ№НцЈ¬ІўХTҢ§үДЛАРФөтНцөД°lЙъЈ¬ұ»ұҠ¶аёЯ·ЦОД«IЗаІAЎЈAbMoleһйИ«ЗтҝЖСРҝН‘фМṩёЯјғ¶ИЎўёЯЙъОп»оРФөДТЦЦЖ„©Ўўјҡ°ыТтЧУЎўИЛФҙҶОҝ№ЎўМмИ»®aОпЎўҹЙ№вИҫБПЎў¶алДЎў°Рьcө°°ЧЎў»ҜәПОпҺмЎўҝ№ЙъЛШөИҝЖСРФҮ„©Ј¬И«ЗтҙуБҝОД«IҢЈАыТэУГЎЈ
Т»ЎўZ-VAD-FMKөДЧчУГҷCАн
јҡ°ыөДөтНцҝЙТФ·ЦһйғИФҙРФУ|°lәННвФҙРФУ|°lғЙ·NоҗРНЈ¬ғИФҙРФНҫҸҪЈәУЙјҡ°ығИІҝӯhҫіЧғ»ҜТэ°lЈ¬Из DNA “pӮыЎўСх»Ҝ‘ӘјӨЎўҫҖБЈуw№ҰДЬ®җіЈөИЎЈҙЛ•rЈ¬ҫҖБЈуwДӨНЁНёРФёДЧғЈ¬бҢ·Ејҡ°ыЙ«ЛШ CЈЁCytochrome CЈ©ЦБјҡ°ыЩ|Ј¬јӨ»оөтНцПакPөД°ллЧМмө°°ЧГёЈЁCaspaseЈ©ЎЈНвФҙРФНҫҸҪЈәНЁЯ^јҡ°ыұнГжЛАНцКЬуwЈЁИз FasЎўTNFR1Ј©ҪУКЬНвІҝРЕМ–ЈЁИзFasLЎўTNF-αЈ©Ј¬РОіЙЛАНцХTҢ§РЕМ–ҸНәПуwЈЁDISCЈ©Ј¬ЦұҪУјӨ»оЖрКјРНCaspaseЈЁИз Caspase-8ЎўCaspase-9Ј©ЎЈ®”өтНцұ»јӨ»оәу•юЯMИлCaspaseјүВ“·ҙ‘ӘЈәјӨ»оөДЖрКјРНCaspaseЯMТ»ІҪЗРёоІўјӨ»оР§‘ӘРН CaspaseЈЁ Caspase-3ЎўCaspase-6ЎўCaspase-7Ј©ЎЈР§‘ӘРН Caspase ЧоәуНЁЯ^ЗРёојҡ°ығИкPжIө°°ЧЈЁИзјҡ°ы№ЗјЬө°°ЧЎўDNA РЮҸНГё PARP өИЈ©Ј¬Тэ°lјҡ°ыҪYҳӢұАҪвәН№ҰДЬҶКК§ЎЈ

ҲD1. јҡ°ыөтНцНЁВ·ҲD[1]
Z-VAD-FMKЈЁZ-VAD(OH)-FMKЈ¬AbMoleЈ¬M3143Ј©ҝЙТФТЦЦЖCaspaseјТЧеЦРөДCaspase-1өҪCaspase-10өИ9·NлЧМмө°°ЧГёөД»оРФЈЁZ-VAD-FMKҢҰCaspase-2өДТЦЦЖДЬБҰУРПЮЈ©ЎЈCaspaseјТЧеФЪҪYҳӢЙПҫЯУРТ»¶ЁөДПаЛЖРФЈәЛщУРөДCaspase¶ј°ьАЁТ»ҙуТ»РЎғЙӮҖҙЯ»ҜҒҶ»щЈ¬УГУЪЗРёоөЧОпө°°ЧЎЈҙЛНвЈ¬Т»Р© Caspaseә¬УРDEDЈЁЛАНцР§‘ӘҪYҳӢУтЈ©»тCARDЈЁCaspaseДјјҜҪYҳӢУт Ј©Ј¬Ш“ШҹCaspaseөД№СҫЫ»ҜәНјӨ»оТФј°ЛАНцХTҢ§РЕМ–ҸНәПОпҪMСbЎЈZ-VAD-FMKЈЁAbMoleЈ¬M3143Ј©НЁЯ^І»ҝЙДжөШҪYәП CaspaseөДҙЯ»ҜО»ьcҒнТЦЦЖЖд»оРФЈ¬ЯM¶шТЦЦЖјҡ°ыөДөтНцЎЈ
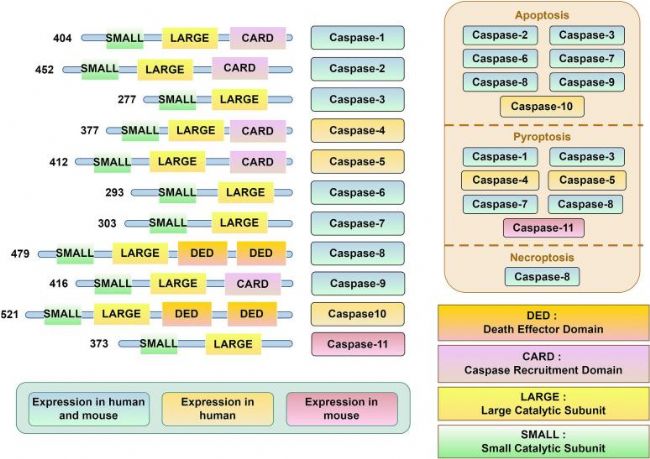
ҲD2. CasepaseјТЧеәН·Цоҗ[2]
2.Z-VAD-FMKТЦЦЖҪ№Нц
јҡ°ыҪ№НцКЗТ»·NУЙCaspaseЈЁИз Caspase-1ЎўCaspase-4ЎўCaspase-5Ј©ҪйҢ§өДҙЩСЧРФјҡ°ыЛАНц·ҪКҪЎЈТ»°гУЙІЎФӯуwПакP·ЦЧУДЈКҪЈЁPAMPsЈ©»т“pӮыПакP·ЦЧУДЈКҪЈЁDAMPsЈ©У|°lСЧ°YРЎуwЈЁИзNLRP3ЎўAIM2Ј©өДҪMСbЈ¬ХРДјІўјӨ»оCaspase-1З°уwЎЈ»о»ҜөДCaspase-1ЗРёоGasdermin DЈЁGSDMDЈ©Ј¬бҢ·ЕЖд N ¶ЛҪYҳӢУтЈ¬Ф“ҪYҳӢУт№СҫЫ»ҜәуҝЙФЪјҡ°ыДӨЙПРОіЙҝЧ¶ҙЈ¬Ң§ЦВјҡ°ығИИЭОпбҢ·ЕІўТэ°lҸҠБТөДСЧ°Y·ҙ‘ӘЎЈZ-VAD-FMKЈЁAbMoleЈ¬M3143Ј©ҝЙТФТЦЦЖCaspase-1ЯM¶шЧи”аСЧ°YРЎуwөДРОіЙЈ¬ЧоҪKҢҚ¬FҢҰјҡ°ыҪ№НцЈЁPyroptosisЈ©өДУРР§ТЦЦЖЎЈ
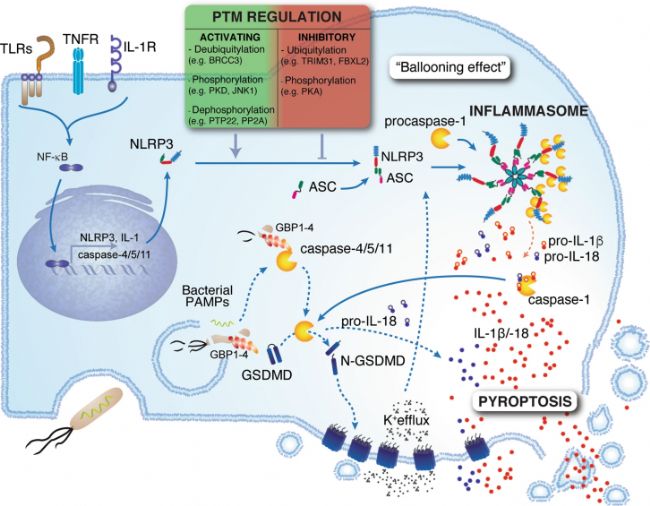
ҲD3. јҡ°ыҪ№НцКҫТвҲD[1]
3.Z-VAD-FMKХTҢ§үДЛАРФөтНц
үДЛАРФөтНцУЙД[БцүДЛАТтЧУЈЁTNFЈ©і¬јТЧеКЬуwЈЁИзTNFR1ЎўFasЈ©»тДЈКҪЧR„eКЬуwЈЁИзTLR3/4Ј©јӨ»оЈ¬КЬуwјӨ»оәуҝЙХРДј RIPK1ЎўCaspase-8ЎўCaspase-10өИРОіЙҸНәПОп IЈЁДӨҪYәПРНЈ©Ј¬ҙЛ•rCaspase-8•юјфЗРRIPK3ТФТЦЦЖүДЛАРЎуwөДРОіЙЈ¬ТтҙЛЙПКцРЕМ–НЁВ·Ж«ПтТэЖрјҡ°ыөтНцЎЈ¶ш®”Caspase-8»оРФұ»ТЦЦЖЈЁИзК№УГ Z-VAD-FMKЈ©•rЈ¬RIPK1ЕcRIPK3ҪYәПРОіЙҸНәПОпIIbЈ¬ТЖО»ЦБјҡ°ыЩ|Ј¬лSәуRIPK3НЁЯ^БЧЛб»ҜЖдөЧОпMLKLЈЁ»мәПЧVПөјӨГёҪYҳӢУтҳУө°°ЧЈ©Ј¬ХTҢ§MLKL№СҫЫ»ҜІўЮDО»ЦБјҡ°ыДӨЈ¬РОіЙҝЧ¶ҙҢ§ЦВјҡ°ыД[ГӣЖЖБСЈ¬бҢ·ЕDAMPsІўТэ°lСЧ°Y·ҙ‘ӘЎЈФЪҫЯуwөДҢҚтһЦРЈ¬Z-VAD-FMKЈЁ ·ъјЧ»щНӘЈ¬AbMoleЈ¬M3143Ј©іЈіЈәНTNF-αЈЁAbMoleЈ¬M9370Ј©В“әПК№УГХTҢ§јҡ°ы°lЙъүДЛАРФөтНц[3]ЎЈ2014ДкЈ¬AbMoleөДғЙҝоТЦЦЖ„©·Ц„eұ»Оч°аСАҮшјТРДСӘ№ЬСРҫҝЦРРДәНГАҮшёзӮҗұИҒҶҙуҢWУГУЪ„УОпуwғИҢҚтһЈ¬ПакPҝЖСРіЙ№ы°lұнУЪн”ҝҜ Nature әН Nature MedicineЎЈ

ҲD4. јҡ°ыүДЛАРФөтНцКҫТвҲD[1]
¶юЎў·¶АэФ”Ҫв
1.Cancer Res. 2024 Apr 1;84(7):1084-1100.
ЙПКцОДХВЦчТӘСРҫҝБЛ»ҜәПОпONC213ФЪјұРФЛиПө°ЧСӘІЎЈЁAMLЈ©ЦРөДЧчУГЎЈСРҫҝ°l¬FЈ¬ONC213НЁЯ^°РПтα-НӘОм¶юЛбГ“ҡдГёЈЁα-KGDHЈ©ҒнХTҢ§ҫҖБЈуw‘ӘјӨәНТЦЦЖСх»ҜБЧЛб»ҜЈ¬ҸД¶шФЪAMLјҡ°ыЦР°l“]ҸҠҙуөДТЦЦЖ»оРФЈ¬УИЖдКЗФЪAMLёЙјҡ°ыЦРЈ¬Н¬•rҢҰХэіЈФмСӘјҡ°ыөД¶ҫРФҳOөНЎЈҙЛНвЈ¬ONC213ЯҖНЁЯ^ТЦЦЖMCL-1өД·ӯЧgҒнҙЩЯMAMLјҡ°ыөДөтНцЎЈҒнЧФAbMoleөДZ-VAD-FMKЈЁAbMoleЈ¬M3143Ј©ұ»УГҒнтһЧCONC213ХTҢ§өДјҡ°ыЛАНцКЗ·сТАЩҮУЪCaspaseҪйҢ§өДөтНцНҫҸҪЈ¬ҪY№ып@КҫZ-VAD-FMKДЬүтп@ЦшХьҫИMV4–11јҡ°ыГвКЬONC213ХTҢ§өДјҡ°ыөтНц[4]ЎЈ
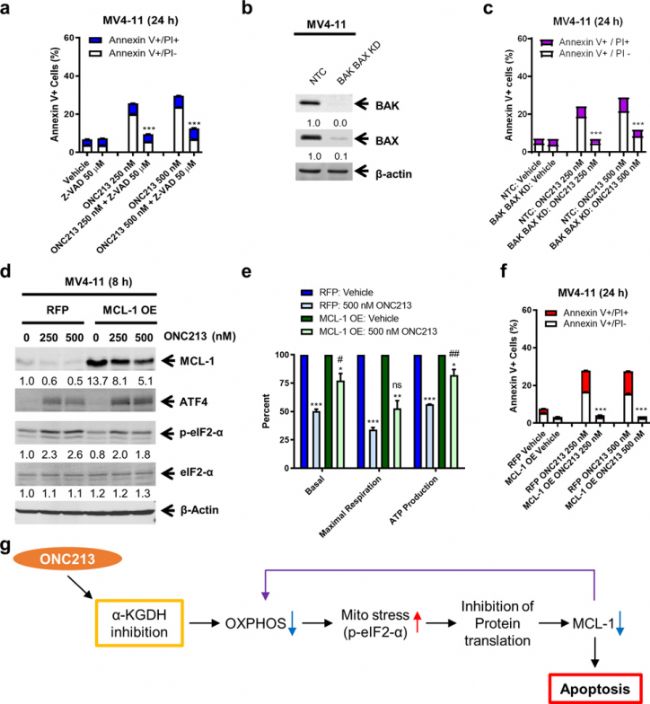
ҲD5. Relationship between MCL-1 and ONC213 sensitivity[4].
2.Cancer Res. 2023 Jan 18;83(2):251-263
ЙПКцОДХВСРҫҝБЛMEX3AФЪВСіІ°©јҡ°ыЦРөДЧчУГЈ¬МШ„eКЗЖдИзәОНЁЯ^Х{№қp53ө°°ЧҒнУ°н‘Д[БцөДЙъйLәНиFЛАНцЎЈСРҫҝ°l¬FЈ¬MEX3AФЪВСіІ°©јҡ°ыЦРЯ^ұнЯ_Ј¬УИЖдКЗФЪә¬УРТ°ЙъРНp53өДВСіІ°©ҒҶРНЦРЎЈЧоҪKҪY№ыұнГчMEX3AНЁЯ^ҙЩЯMp53ө°°ЧөДҪөҪвЈ¬ТЦЦЖБЛp53ҪйҢ§өДиFЛАНцЈ¬ҸД¶шҙЩЯMБЛД[БцөДЙъйLәНҙж»оЎЈФЪјҡ°ыҢҚтһЦРСРҫҝИЛҶTҢҰMEX3AИұК§ЈЁsh-MEX3AЈ©әНҢҰХХЈЁsh-CtrlЈ©өДPA-1әНTOV21Gјҡ°ыЯMРРБЛІ»Н¬ТЦЦЖ„©өДМҺАнЈ¬ТФҙ_¶ЁMEX3AИұК§ФміЙөДјҡ°ыЛАНцоҗРНЎЈФ“Іҝ·ЦөДҢҚтһК№УГБЛҒнЧФAbMoleөДZ-VAD-FMK (өтНцТЦЦЖ„©Ј¬AbMoleЈ¬M3143)Ј¬Necrostatin-1 (үДЛАТЦЦЖ„©Ј¬AbMoleЈ¬M2315)әН3-Methyladenine (ЧФКЙТЦЦЖ„©Ј¬AbMoleЈ¬M2296)өИ¶аҝо®aЖ·[5]ЎЈ
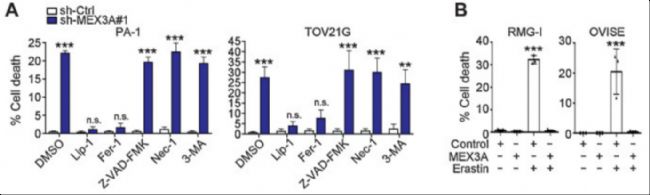
ҲD6. MEX3A depletion leads to ferroptosis phenotypes in WT p53 ovarian cancer cells[5]
3.J Hazard Mater. 2021 Mar 15;406:124682.
ЙПКцОДХВөДәЛРДДҝөДКЗСРҫҝБт»ҜҡдЈЁH2SЈ©ҢҰИвлu·ЁКПДТЈЁBursa of FabriciusЈ©өДГвТЯ“pӮыҷCЦЖЈ¬ҪY№ыЧCҢҚH2Sұ©В¶ҝЙҢ§·ЁКПДТјҡ°ыөДүДЛАРФөтНцәНСЧ°Y·ҙ‘ӘЈ¬®aЙъСх»Ҝ‘ӘјӨЈ¬јӨ»оTGF-βРЕМ–НЁВ·Ј¬ЙПКцЯ@Р©Я^іМУЙmiR-15b-5p/TGFBR3ЭS№ІН¬Х{№қЎЈФЪҢҚтһЦРЈ¬ҝЖСРИЛҶTК№УГБЛУЙAbMoleМṩөДZ-VAD-FMKЈЁAbMoleЈ¬M3143Ј©ЎўNecrostatin-1ЈЁAbMoleЈ¬M2315Ј©ТФтһЧCүДЛАРФөтНцФЪЙПКцЯ^іМЦРөДЧчУГ[6]ЎЈ
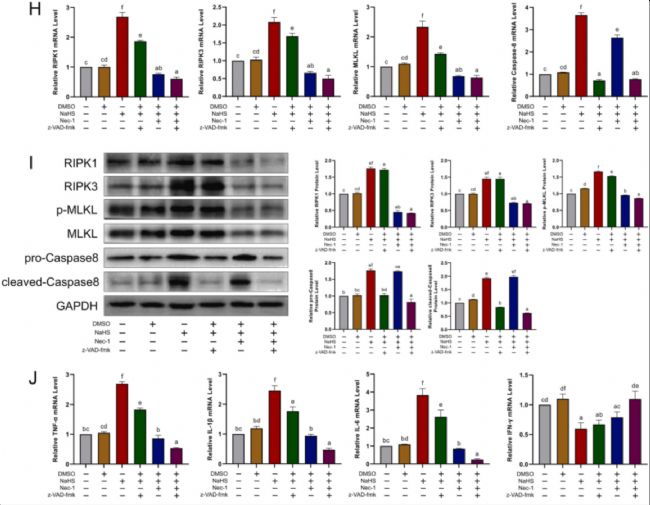
ҲD7. NaHS treatment induces necroptosis and inflammation in DT40 and HD11[6]

…ўҝјОД«Iј°шQЦx
[1] Damien Bertheloot, Eicke Latz, Bernardo S. Franklin, Necroptosis, pyroptosis and apoptosis: an intricate game of cell death, Cellular & Molecular Immunology 18(5) (2021) 1106-1121.
[2] W. Zhang, H. Wu, Y. Liao, et al., Caspase family in autoimmune diseases, Autoimmunity reviews 24(2) (2025) 103714.
[3] C. Yu, Z. Lei, X. Li, et al., Role of HMGB1 in TNF-α Combined with Z-VAD-fmk-Induced L929 Cells Necroptosis, Biochemical genetics 60(2) (2022) 598-610.
[4] Y. Su, J. L. Carter, X. Li, et al., The Imipridone ONC213 Targets α-Ketoglutarate Dehydrogenase to Induce Mitochondrial Stress and Suppress Oxidative Phosphorylation in Acute Myeloid Leukemia, Cancer research 84(7) (2024) 1084-1100.
[5] C. K. Wang, T. J. Chen, G. Y. T. Tan, et al., MEX3A Mediates p53 Degradation to Suppress Ferroptosis and Facilitate Ovarian Cancer Tumorigenesis, Cancer research 83(2) (2023) 251-263.
[6] C. Qianru, H. Xueyuan, Z. Bing, et al., Regulation of H(2)S-induced necroptosis and inflammation in broiler bursa of Fabricius by the miR-15b-5p/TGFBR3 axis and the involvement of oxidative stress in this process, Journal of hazardous materials 406 (2021) 124682.
ҳЛәһЈә
ТЦЦЖ„©

- Mitomycin CөДЧчУГҷCЦЖј°ФЪД[Бцјҡ°ыПөәН„УОпДЈРНЦРөД¶аҲцҫ°‘ӘУГ
- CFDA-SEөДЧчУГҷCЦЖј°ФЪјҡ°ыФцЦіәНуwғИКҫЫҷЦРөД‘ӘУГ
- ТЦЦЖ„©A83-01өДЧчУГҷCЦЖј°ФЪёЙјҡ°ыЕcоҗЖч№ЩЕарBЦРөД‘ӘУГ
- 2-Deoxy-D-glucoseЈЁ2-DGЈ©өДЧчУГҷCЦЖј°ФЪМЗҪНҪвТЦЦЖСРҫҝЦРөДЧчУГ
- в}лxЧУҹЙ№вМҪбҳCalbryte 520.AMФЪ„У‘BұOңyЦРөДәЛРДғһ„ЭЕc‘ӘУГҪвОц
- NRF2әНAKTјӨ»о»тFBP1ИұК§ФЪёОјҡ°ыЛҘАПәНёО°©ЯMХ№ЦРөДЧчУГ
- ¶алДөД№ҰДЬәН‘ӘУГј°Жд№МПаәПіЙјјРg
- иFЛАНцТЦЦЖ„©Ferrostatin-1өДЧчУГҷCЦЖЕc‘ӘУГ
- MCEБщФВХ№•юоAёжЈәИЛ№ӨЦЗДЬҙу•ю&ёЙјҡ°ыСРҫҝ•юЧh
- MCE HOLOПөБРҫщПаГвТЯ·ЦОцФҮ„©әРЙПРВ5ХЫҮLхrғr
- MCEЦұІҘоAёжЈәPROTACСРҫҝРВЯMХ№әНФOУӢРВЛјВ·
- MCE¶ЛОзлpЦШёЈАыЈәКЦЛЩPKЩҗәНҹбйT¶алД®aЖ·қMЩӣ¶YЖ·
- MCEЦРҮшіхПДҙЩдN»о„УЈәИҫБП®aЖ·в »Э55ХЫҝсҡg
- MCEЕcДгПајsөЪБщҢГҮшлHјҡ°ыЕc»щТтЦОҜҹЦРҮш·е•ю
- Оч°І°ЩОһ618ДкЦРҙуҙЩ,ЩIҹЙ№вИҫБПЛНёЯНЁБҝҢЈУГОўҝЧ°е
- ОчГАҪЬіЙ№Ұ…ўјУөЪ¶юК®ЛДҢГЦРҮшЙъОпЦЖЖ·ҙу•ю


Copyright(C) 1998-2025 ЙъОпЖчІДҫW лҠФ’Јә021-64166852;13621656896 E-mailЈәinfo@bio-equip.com