脂肽介導CRISPR-Cas9遞送基因編輯在遺傳性眼病治療新方法中的應用
Stargardt 病(STGD1)是一種由 ABCA4 基因的雙等位基因變異引起的遺傳性黃斑變性疾病。ABCA4 基因編碼一種定位在光感受器外段的轉運蛋白,負責清除視覺循環中的有毒副產物。目前已發現超過 1,200 種 STGD1 致病性變異,其中約 10% 是隱藏在基因深處的“深內含子(deep-intronic, DI)突變”——它們會讓細胞在讀取基因時錯誤插入多余的“偽外顯子”,導致生成的蛋白質無法正常工作或被提前降解。目前,在研的反義寡核苷酸療法需要為每個突變單獨設計序列,適用范圍很小。因此,開發一種糾正不同位點突變引起的剪接缺陷的通用療法具有重要意義。
近日,荷蘭內梅亨大學醫學中心和烏特勒支大學藥物科學研究所的科研團隊在 Molecular Therapy – Nucleic Acids 雜志上聯合發表了一項突破性研究。他們利用 CRISPR-Cas9 基因編輯系統,結合兩組導向 RNA(gRNA),分別靶向 ABCA4 基因中兩個高頻 DI 變異區域(內含子 30 和 36),并通過脂肽 C18:1-LAH5 實現 Cas9 核糖核蛋白(Cas9 ribonucleoproteins, Cas9 RNP)的高效遞送。該策略不僅能精準切除錯誤插入的基因片段,還能保留正常蛋白功能。該策略在視網膜前體細胞中的成功應用,為開發通用型基因療法奠定了基礎。這項突破不僅為 Stargardt 病患者帶來新希望,也為其他遺傳病的治療開辟了新思路。

一、RPNCs 綜合表征
研究者分別設計了靶向 ABCA4 基因內含子 30 和 36 的 gRNA,并選擇了油酸修飾的 LAH5 多肽(即脂肽 C18:1-LAH5)作為遞送載體。C18:1-LAH5 與 Cas9 RNP 孵育后形成納米復合物(ribonucleoprotein nanocomplexes, RPNCs),通過動態光散射(DLS)監測 RPNC 的形成。結果顯示,隨著 Cas9 RNP: 脂肽比例提高,RPNCs 的粒徑減小,平均粒徑在 151-191 nm 之間;隨著脂肽濃度的增加,RPNCs 的 Zeta 電位逐步升高(圖 1A),此外,電泳遷移率轉移試驗(electrophoretic mobility shift assay, EMSA)顯示(圖 1B),RPNCs 的遷移率隨著脂肽濃度的增加顯著降低,在 GFP-Cas9:ATTO550-gRNA:ATTO647-gRNA:脂肽=1:1:1:50 的比例下即可合成 RPNCs 穩定產物。綜上結果表明,脂肽 C18:1-LAH5 能夠與 Cas9 RNP 高效結合,形成穩定的納米復合物。
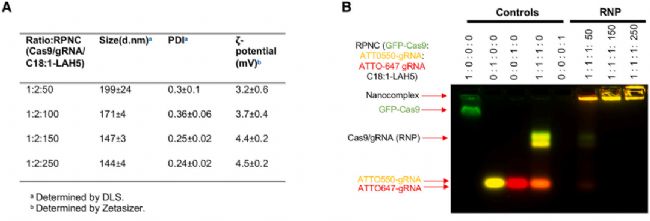
圖1. RNP 和 C18:1-LAH5 脂肽不同比例下合成的 RPNCs 表征
利用 NanoFCM 在單顆粒水平對 RPNCs 進行綜合表征,檢測參數包括顆粒粒徑、濃度和包封率等多個維度。結果顯示,與不含 C18:1-LAH5 脂肽的樣品相比,Cas9 RNP 與 C18:1-LAH5 脂肽形成 RPNCs 后(無論是單個 gRNA 還是兩個 gRNA 的組合)中位直徑和熒光陽性顆粒的濃度均顯著增加(圖 2A-D);此外,RPNCs 在單顆粒水平兩種 gRNA 雙陽的比例高達 84%,比不含脂肽的樣品比例高出 70 倍(圖 2E-F)。綜上表明,C18:1-LAH5 脂肽能夠有效促進復合物形成 RPNCs,而不含 C18:1-LAH5 脂肽的樣本中熒光顆粒很可能是 Cas9 RNP 的聚集物。
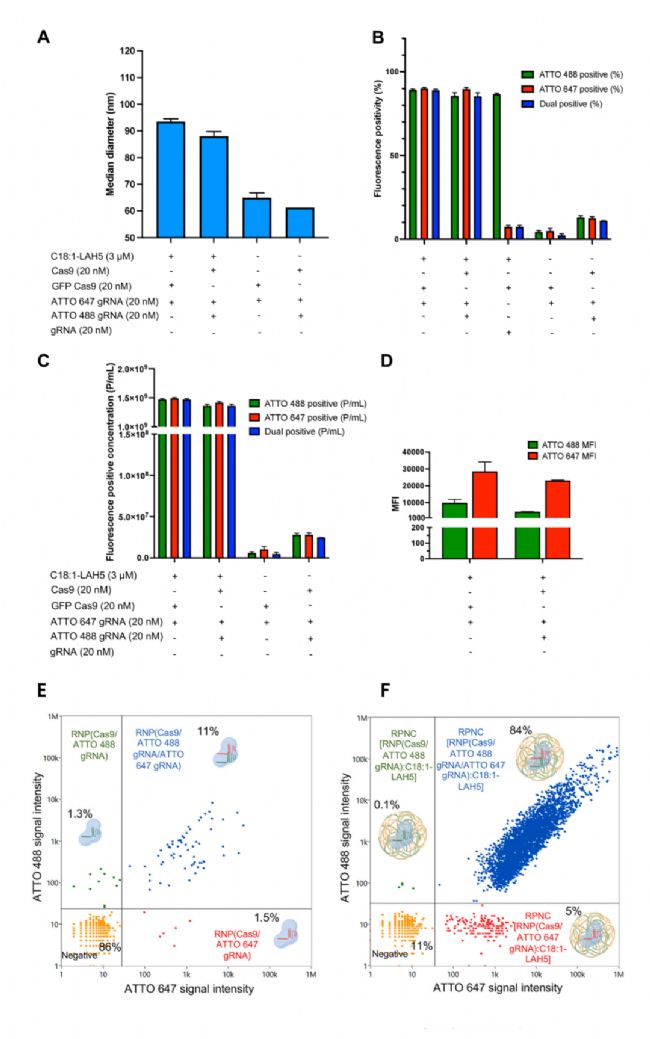
圖2. NanoFCM 對 RPNCs 的綜合表征
二、細胞攝取效率
為了觀察細胞對 RPNCs 的攝取,在 HeLa 細胞中加入 RPNCs,并在分別培養 2、4、12、24 小時后進行熒光共聚焦成像。結果顯示,隨培養時間的延長,GFP-Cas9、ATTO550-gRNA1和ATTO647-gRNA2 的熒光信號均逐步增強,表明 HeLa 細胞能同時攝取 RPNCs 中的 Cas9 和兩種 gRNA(圖 3)。

圖3. 共聚焦熒光顯微鏡下細胞攝取 RPNCs
三、基因編輯效率
明確了細胞對 RPNCs 的有效攝取后,為了驗證 RPNCs 模型的基因編輯能力,研究者們選用了 HEK293T stoplight 和 eGFP HEPA 兩種熒光報告細胞系對 RPNCs 的編輯效率進行評估。首先在 HEK293T stoplight 細胞系中分別使用脂肽 C18:1-LAH5 和 LAH5 修飾的 RPNCs 進行 eGFP 的插入,如圖 4A、B 所示,脂肽 C18:1-LAH5 的組別中 eGFP 的表達量顯著高于 LAH5;而后在 eGFP HEPA 細胞中進行 eGFP 敲除,如圖 4C、D 所示,脂肽 C18:1-LAH5 的組別敲除比例顯著高于 LAH5,且隨濃度升高而升高。綜上表明 RPNCs 能夠有效地對目標片段進行插入和敲除,而脂肽 C18:1-LAH5 的遞送效率顯著高于 LAH5,且 RNP 與脂肽比例為 1:250 時均呈現較高的編輯效率。

圖4. 兩種熒光報告細胞系中 RPNCs 的基因編輯效率
在明確 RPNCs 的最優合成比例、遞送條件后,研究者們在 HeLa 細胞、患者來源的成纖維細胞、誘導多能干細胞衍生的光感受器前體細胞(PPCs)等不同來源細胞系中進行了細胞攝取實驗,探究了 RPNCs 的基因編輯效率,例如在成纖維細胞模型中,針對內含子 30,gRNAs(30 1+2)在對照組和 STGD1 患者組中均實現了部分內含子移除,編輯效率均在 75% 左右,顯著高于 gRNAs(30 3+4)(圖 5A)。針對內含子 36,兩對 gRNA(gRNAs 36 1+2和gRNAs 36 1+3)都能誘導部分缺失,效率均為 40% 左右(圖 5B)。此外,研究者們還在基因組水平、RNA 水平和蛋白質水平對 RPNCs 的基因剪切、剪切后修復等方面進行了研究,證明了脂肽 C18:1-LAH5 修飾的 RPNCs 在不同細胞模型中均表現出優秀的基因編輯效率,為 STGD1 的治療提供了有力支持。
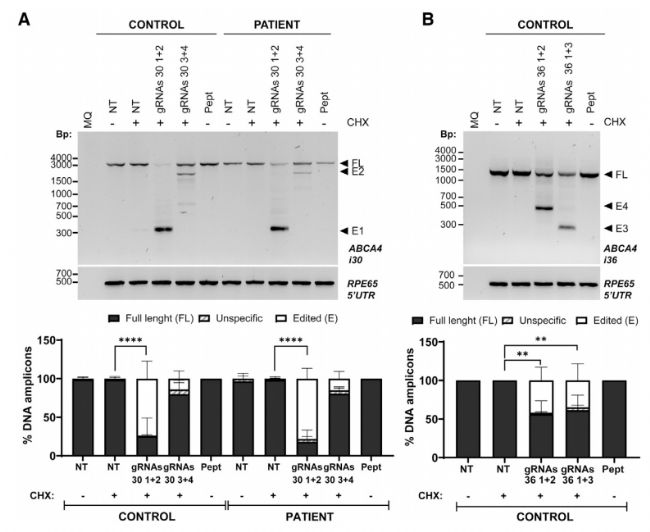
圖5. 成纖維細胞中 RPNCs 的基因編輯效率
研究意義
該研究提出了一種基于脂肽介導 CRISPR-Cas9 遞送的通用基因編輯策略,能夠糾正 ABCA4 基因中由 DI 突變引起的剪接缺陷。這種方法不僅適用于特定突變,還可以同時糾正多個突變,為 STGD1 的治療提供了一種潛在的通用解決方案。此外,脂肽遞送還為非病毒遞送系統在基因編輯中的應用提供了新的思路和方法,推動了基因編輯技術在眼科領域的應用。
展望
NanoFCM 能在單顆粒水平對納米級顆粒的物理化學性質(如粒徑、熒光強度等)進行綜合表征。在基因編輯領域,NanoFCM 可用于表征 RPNCs 的粒徑分布、包封效率、熒光強度等多維信息,這有助于深入理解 RPNCs 的理化性質,為遞送系統的研發和優化提供有力支持。展望未來,隨著基因編輯技術的持續發展,對載體質量、荷載效率等關鍵指標的檢測勢必會成為常規要求,NanoFCM 有望成為基因編輯載體質量控制和穩定性評估等指標的關鍵表征工具,為臨床科研和產業化提供便捷且精準的檢測方式,推動新型治療技術的發展!