
¯ÔĪxĩÄÉúÃüÖÜÆÚē͸ĐČžÉúÎīCÖÆ
¯ŧ˛ĘĮŌģˇNÎÃÃŊŧ˛˛ĄŖŦ¸ĐČžÕßͨŗŖūŗöŦF°lũĄĸ°lĀäēÍÁ÷¸ĐĶŧ˛˛ĄĄŖČįšû˛ģŧ°rÖίŖŦĀÖØÕßÉõÖÁūÎŖŧ°ÉúÃüĄŖĘĀĐlŊMŋĐÂŊü°l˛ŧĩÄĩūąíÃ÷ŖŦ2019 ÄęČĢĮōšĀĶ°lÉú 2.29 |¯ŧ˛˛ĄĀũŖŦËĀĶÚÔ˛ĄĩÄČËĩŗŦß^ 40 ČfĀũĄŖ
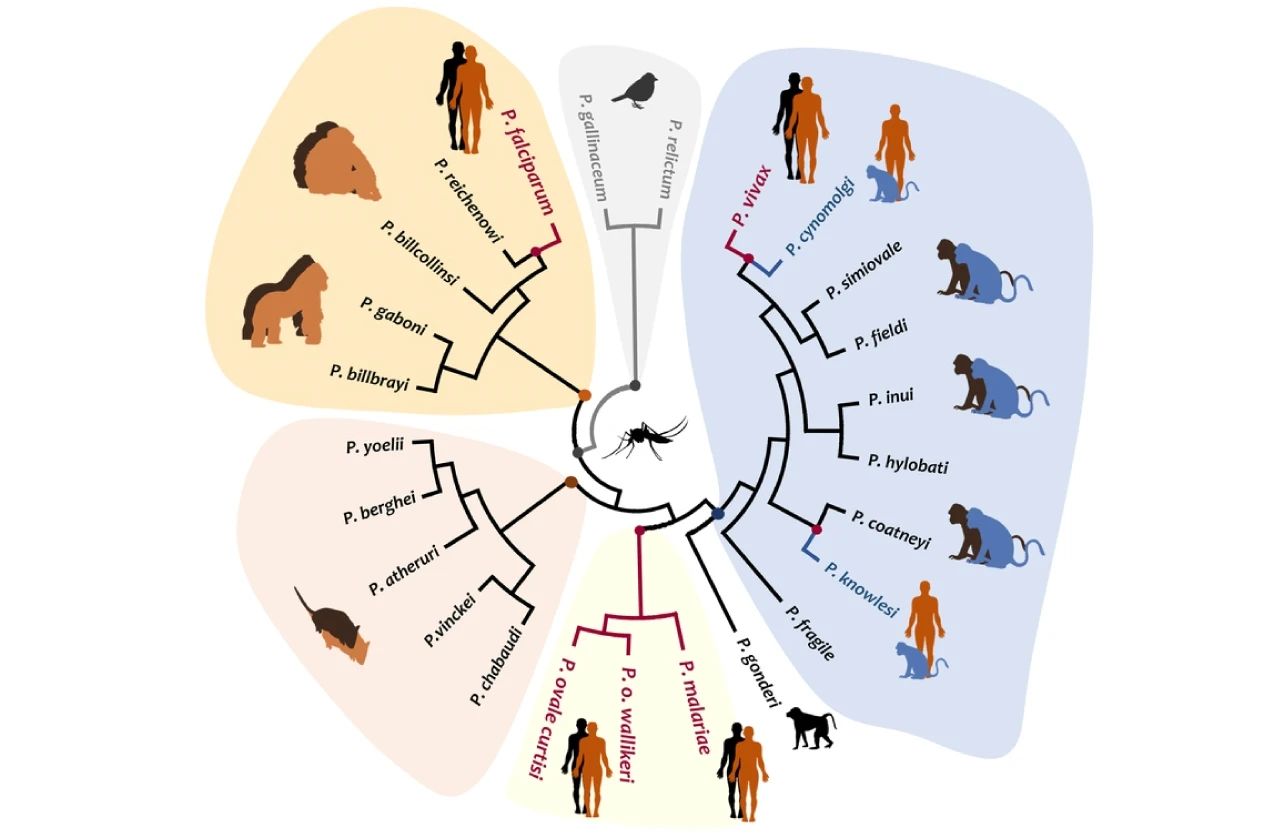
¯ŧ˛ĘĮĶɸĐČžČËîĩÄ 5 ˇNŌŅÖĒ¯ÔĪxÖĐĩÄČÎēÎŌģˇNŌũÆđĩÄÔÉúĶÎīŧ˛˛ĄŖēēĐÔ¯ÔĪxĄĸégČÕ¯ÔĪxĄĸČũČÕ¯ÔĪxĄĸÂŅĐίÔĪxĄĸÖZĘĪ¯ÔĪxĄŖÆäÖĐŖŦÔÚÃĀÖŪēÍWÖŪ×îŗŖŌĩÄĘĮégČÕ¯ÔĪxēÍČũČÕ¯ÔĪxŖŦÔÚˇĮÖŪĘĮēĐÔ¯ÔĪxĄŖ
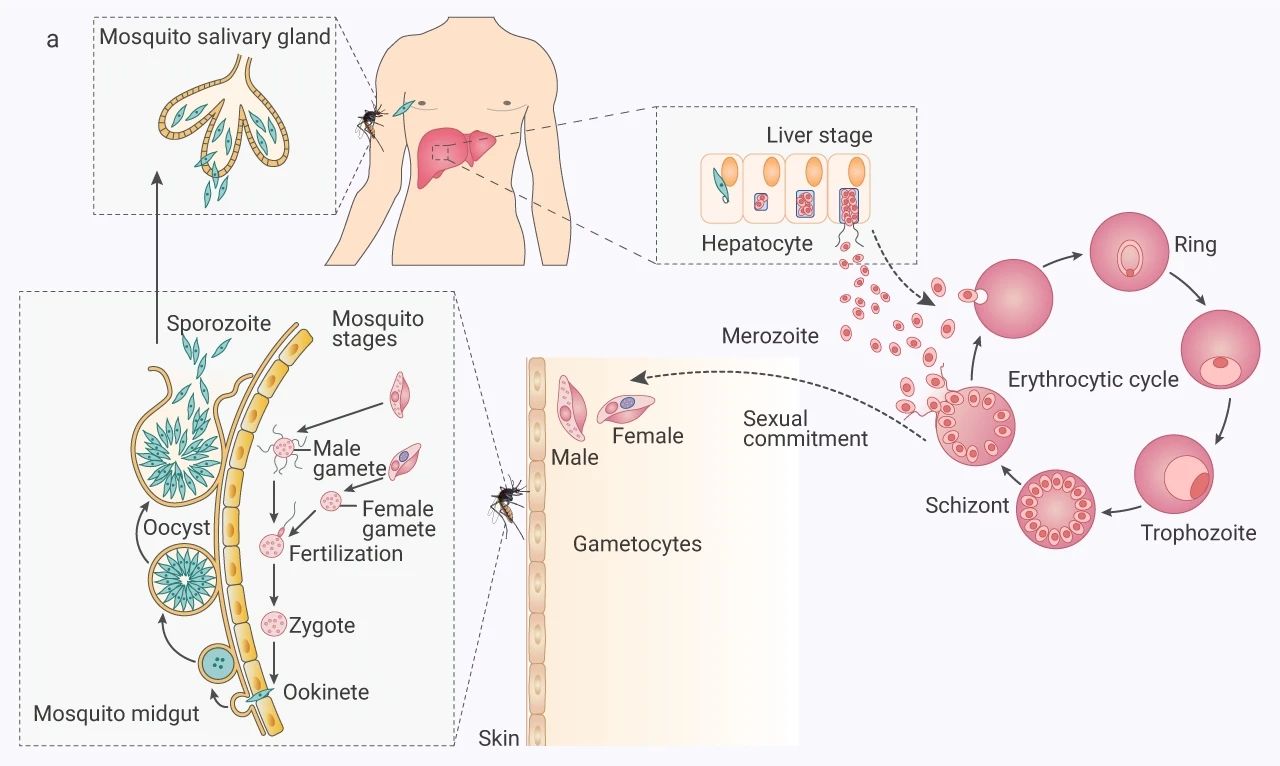
¯ÔĪxÉúÃüÖÜÆÚÖ÷ŌĒˇÖéÉëAļÎŖēŧ´°lÉúČËîËŪÖ÷ČĩÄoĐÔˇąÖŗēÍ°lÉúÔÚÎÃĪxÃŊŊéČĩÄĶĐĐÔˇąÖŗĄŖ
oĐÔˇąÖŗëAļÎŖē´ÆĐÔ°´ÎÃĸēŦĶĐ×Ķæß×Ķ (sporozoites) ĩÄÍŲŌē×ĸČëĩŊČËîËŪÖ÷ķwČĄŖÔÚŌģĐĄrČŖŦß@ĐŠ×Ķæß×ĶßMČëŅĒÁ÷˛ĸŅhĩŊ¸ÎÅKŖŦßMļø¸ĐČž¸Îŧ°û (¸Îŧ°ûrÆÚ)ŖŦÔÚ¸Îŧ°ûČŖŦÃŋ×Ķæß×ĶoĐÔÍÖÆĐÎŗÉĩ°ŲÁŅÖŗ×ĶĄŖ¸Îŧ°ûÆÆÁŅŖŦÁŅÖŗ×ĶáˇÅĩŊŅĒŌēÖĐŖŦĮÖČëŧtŧ°û (ŧtŧ°ûÖÜÆÚ)ĄŖ
ÁŅÖŗ×ĶŊvÔįÆÚh î×ĖđBķw (ring)ĄĸÍíÆÚ×ĖđBķw (trophozoite)ŖŦ×´ÎĶĐŊzˇÖÁŅĐÎŗÉÁŅÖŗķw (schizont) ëAļÎĄŖČģēķÁŅÖŗķwÆÆÁŅŖŦĸÁŅÖŗ×ĶáˇÅģØŅĒŌēÖĐŖŦÔÚÄĮĀīËüŋÉŌÔĮÖČëĐÂĩÄŧtŧ°û˛ĸ ĸÅcŧtŧ°ûŅhĄŖ
ĶĐĐÔˇąÖŗëAļÎ: ÉŲĩ×ĖđBķwŪDģ¯éĐÛĐÔēÍ´ÆĐÔÅä×Ķķw (male/female gametocytes)ĄŖŽ´ÆĐÔ°´ÎÃÎüĘŗĘܸĐČžËŪÖ÷ĩÄŅĒŌērŖŦËüūzČĄß@ĐŠÅä×ĶķwŖŦß@ĐŠÅä×ĶķwÔÚÎÃ×ĶķwČßMĐĐĶĐĐÔŅhŖŦĐÎŗÉ×Ķæß×ĶŖŦÍęŗɯÔĪxĩÄÉúÃüÖÜÆÚĄŖ
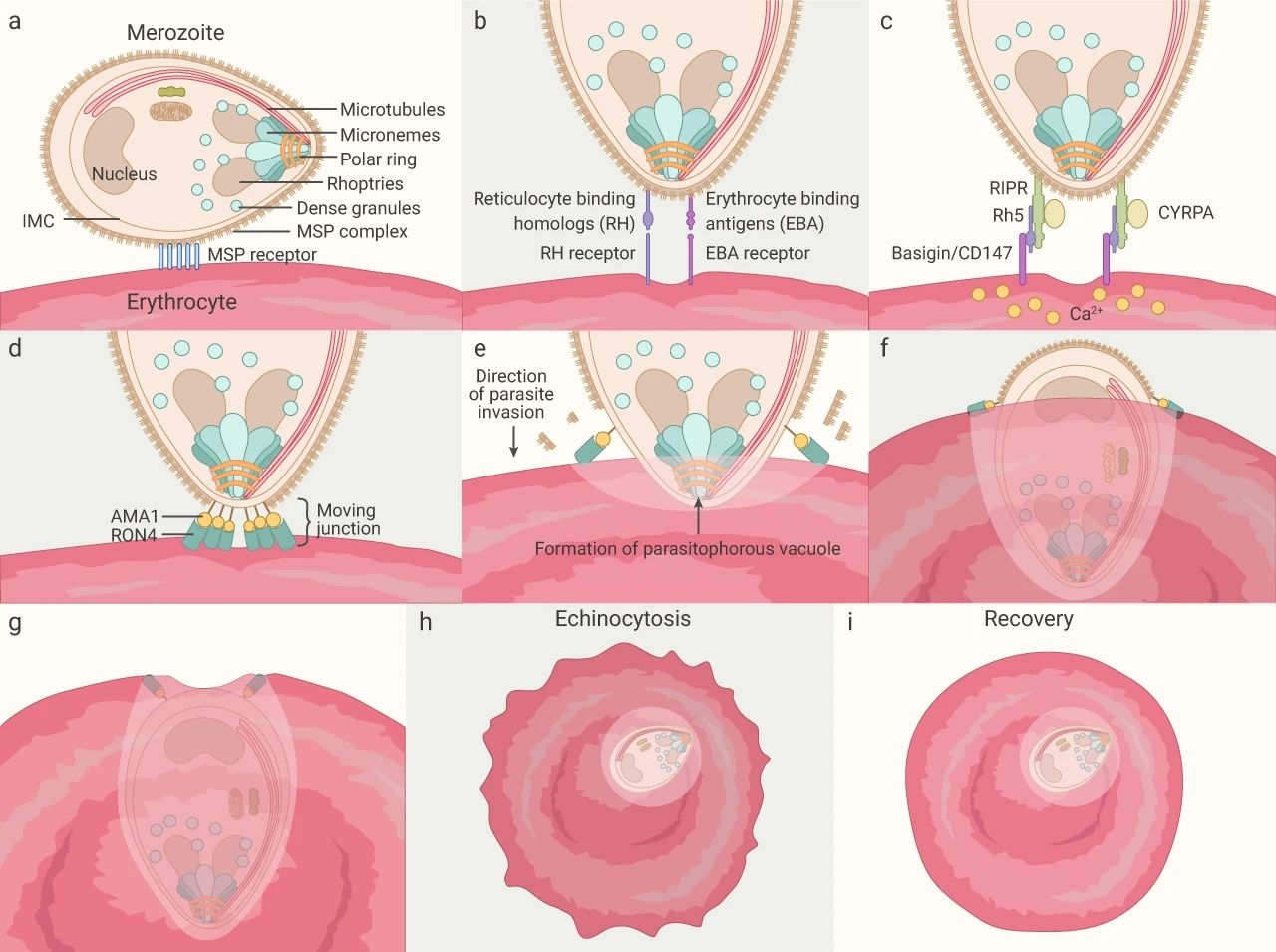
ŌũÆđ°Y îĐÔ¯ŧ˛ĩÄëAļÎÖ÷ŌĒ°lÉúÔÚ¯ÔĪx¸ĐČžĩÄŧtŧ°ûrÆÚŖŦÔÚ´Ë×ößMŌģ˛ŊĩÄŊéŊBĄŖ
ČëĮÖŖē ÁŅÖŗ×ĶēÍŧtŧ°ûÖŽégĩÄŗõĘŧŊĶĶ|ĘĮëSCĩÄŖŦĩĢéÁËŗÉšĻČëĮÖŖŦÁŅÖŗ×ĶĘšĶÃŧĄĶĩ°°×-ŧĄĮōĩ°°×ß\ĶÖØĐÂļ¨ĪōŖŦĘšÆäíļËÅcŧtŧ°ûĤŊĶĶ|ĄŖÁŅÖŗ×ĶÅcŧtŧ°ûĩÄŗõĘŧĪāģĨ×÷ĶÃÉæŧ°ĩŊÁŅÖŗ×ĶąíÃæĩ°°× 1 (MSP1)ŖŦļøÖØĐÂļ¨ĪōēķĩÄíļ˸ŊÖøÉæŧ°ŧÄÉúĪxĖØļ¨ČëĮÖÅäķw (Čį EBA ēÍ PfRh ŧŌ×åŗÉT) ēÍŧtŧ°ûąíÃæĘÜķwĪāģĨ×÷ĶÃĄŖ
ŗũ´ËÖŽÍâŖŦÁŅÖŗ×ĶēÍŧtŧ°ûĩÄžoÃÜßBŊĶŖŦÅc¯ÔĪxÎĸÁŖĩ°°× AMA1 ēÍŧÄÉúĪxÁâĐÎîi (RON) ĩ°°×ĐÎŗÉĩÄÍēĪÎīŌ˛ĶĐęPĄŖ
ÍÖÆŖēŌģĩŠŧtŧ°û¸ĐČžąģŊ¨ÁĸŖŦŧÄÉúĪxŅ¸ËŲÍÖÆŖŦÔÚëSēķĩÄ 48 ĐĄrČŖŦÁŅÖŗķwˇÖÁŅŖŦŽaÉúÁŅÖŗ×ĶŖŦß@ĐŠÁŅÖŗ×ĶÔÚ°lĶũrŌŨŗöŖŦ§ÖÂŧtŧ°ûĤÆÆÄēÍŧÄÉúĪxąŦ°lĐÔáˇÅŖŦ˛ĸČëĮÖĐÂĩÄËŪÖ÷ŧ°ûĄŖ
ĶÉĶÚŧÄÉúĪxÔÚŅ¸ËŲÍÖÆß^ŗĖÖĐĐčŌĒŗÖĀmĩÄēËÜÕËáėíēĪŗÉ DNA ēÍ RNAŖŦŌō´ËŖŦÔSļāŋš¯Ë°ĐĪōā×ā¤ÉúÎīēĪŗÉĄŖČį°ĐĪōļūäČ~ËáßÔø (DHFR) ĩÄ PyrimethamineĄĸCycloguanilŖŦ×čāā×ā¤ÉúÎīēĪŗÉĩÄ AtovaquoneŖŦŌÔŧ°Í¨ß^°ĐĪō DHODH í×čāā×ā¤ĩÄÉúÎīēĪŗÉĩÄ DSM265ĄŖ
ÁíÍâŖŦŧÄÉúĪxĻ IđBÎüĘÕĩÄĘÜpÅc Na+ ĩÄĶĐēĻˇeĀÛĶĐęPŖŦļøĩÍĩČÉúÎīČįŧÄÉúĪxĩÄŲ|Ĥ Na+ ŨŗöąÃ (PfATP4)ŖŦŋÉŌÔŧ°rąÃŗöˇeĀÛĩÄ Na+ŖŦÄļøžSŗÖ Na+ ˇBĄŖÁíÍâŖŦ¯ÔĪx PI4K (ČįPfPI4K) ͨß^´ßģ¯Á×ÖŦõŖŧĄ´ŧ (PI) ĩÄÁ×ËáĸĶĪÂĶÎŧÂͨ¡ŖŦÕ{šļāˇNÉúÎīß^ŗĖŖŦ°üĀ¨ÔÚÁŅÖŗ×Ķ°lĶũß^ŗĖÖĐĩÄĐÅĖŪD§ĄĸĤß\ŨēÍ°ûŲ|ˇÖÁŅĄŖ
ÖĩĩÃŌģĖáĩÄĘĮŖŦÄŋĮ°´ķ˛ŋˇÖŋš¯ËÎīļŧĘĮ°ĐĪōŅĒŌēëAļÎŖŦČį Artemisinin ŧ°ÆäŅÜÉúÎīĩČļŧĘĮĶĐЧĩÄŋš¯ËÎīĄŖ

°l˛ĄŖēÖ÷ŌĒĩÄÖ²ĄCÖÆĘĮ¯ÔĪx¸ĐČžēķŖŦŧtŧ°û°lÉúČÜŅĒŖŦáˇÅŗöŧÄÉúĪxēͯŧ˛ČļžËØ (ąģÕJéĘĮ¯ÔĪxēÍŧÄÉúĪx DNA ĩÄÍēĪÎī)ŖŦßMļøŧ¤ģî TLR9 Ķ|°lŅ×°Yˇ´ĒĄŖ´ËÍâŖŦĘܸĐČžĩÄŧtŧ°ûĩÄĤ×Ķ˛ļøĘʧ×ĐÎÄÜÁĻŖŦß@ˇNÄÜÁĻĩÄĘʧ§ÖÂÃĢŧŅĒšÜ×čČûŖŦŽÖØŌĒÆ÷šŲĘÜĩŊĶ°írŖŦĀÖØrūÎŖŧ°ÉúÃüĄŖ
ŋŊYŖē
žÍÄŋĮ°ļøŅÔŖŦ¯ŧ˛ČÔČģĘĮČĢĮōÖØ´ķĩÄŧ˛˛ĄØúĄŖąMšÜ Artemisinin ŧ°ÆäŅÜÉúÎīĘĮŌģžŋš¯ËÎīŖŦArtemisinin ÂēĪ¯ˇ¨Đ§šû¸üĘĮĘŽˇÖī@ÖøŖŦĩĢÄÍËĐÔŧÄÉúĪxĩÄŗöŦFČÔÅfĘĮÖØŌĒĩÄî}ĄŖŌō´ËŖŦĐčŌĒé_°lĐ°ĐücĩÄĐÂĐÍŋš¯ËŖŦČįģųĶÚŧtŧ°ûČëĮÖĐčŌĒŌģĪĩÁĐ fÕ{ĩÄĩ°°×Ų|/ĩ°°×Ų|ĪāģĨ×÷ĶÃĩČĄŖÁíÍâŖŦÅcŧtŧ°ûąíÃæĩÄ basigin ĪāģĨ×÷ĶÃĩÄēĐÔ¯ÔĪxĩ°°×ÍēĪÎī PfRH5ĄĸPfAMA1 ĩČŌ˛ŋÉ×÷éŌßÃįŅĐžŋĄŖŌĒØĩ×ĪûŗũČËî¯ŧ˛ŖŦÎŌČÔĶĐŌģļΡŌĒ×ߥŖ
MCE ĩÄËųĶĐŽaÆˇHĶÃ×÷ŋÆWŅĐžŋģōË×CÉęķŖŦÎŌ˛ģéČÎēÎČËĶÃÍžĖᚊŽaÆˇē͡ūÕ
ĸŋŧÎÄĢI
1. Phillips MA, et al. Malaria. Nat Rev Dis Primers. 2017;3:17050.
2. Escalante AA, et al. Malaria Molecular Epidemiology: An Evolutionary Genetics Perspective. Microbiol Spectr. 2019;7(4):10.1128/microbiolspec. AME-0010-2019.
3. De Koning-Ward TF, et al. Plasmodium species: master renovators of their host cells. Nat Rev Microbiol. 2016;14(8):494-507.
4. Zheng Z, et al. Advances in Molecular Diagnosis of Malaria. Adv Clin Chem. 2017;80:155-192.
5. Cowman AF, et al. Malaria: Biology and Disease. Cell. 2016;167(3):610-624.
6. Mohandas N, et al. Malaria and human red blood cells. Med Microbiol Immunol. 2012;201(4):593-598.
7. Liang X, et al. Discovery of 6'-chloro-N-methyl-5'-(phenylsulfonamido)-[3,3'-bipyridine]-5-carboxamide (CHMFL-PI4K-127) as a novel Plasmodium falciparum PI(4)K inhibitor with potent antimalarial activity against both blood and liver stages of Plasmodium. Eur J Med Chem. 2020;188:112012.
8. Hisaeda H, et al. Malaria: immune evasion by parasites. Int J Biochem Cell Biol. 2005;37(4):700-706.
9. Osii RS, et al. The Impact of Malaria Parasites on Dendritic Cell-T Cell Interaction. Front Immunol. 2020;11:1597. Published 2020 Jul 24.
10. Yang J, et al. Advances in the research on the targets of anti-malaria actions of artemisinin. Pharmacol Ther. 2020;216:107697.
11. Delves M, et al. The activities of current antimalarial drugs on the life cycle stages of Plasmodium: a comparative study with human and rodent parasites. PLoS Med. 2012;9(2):e1001169.
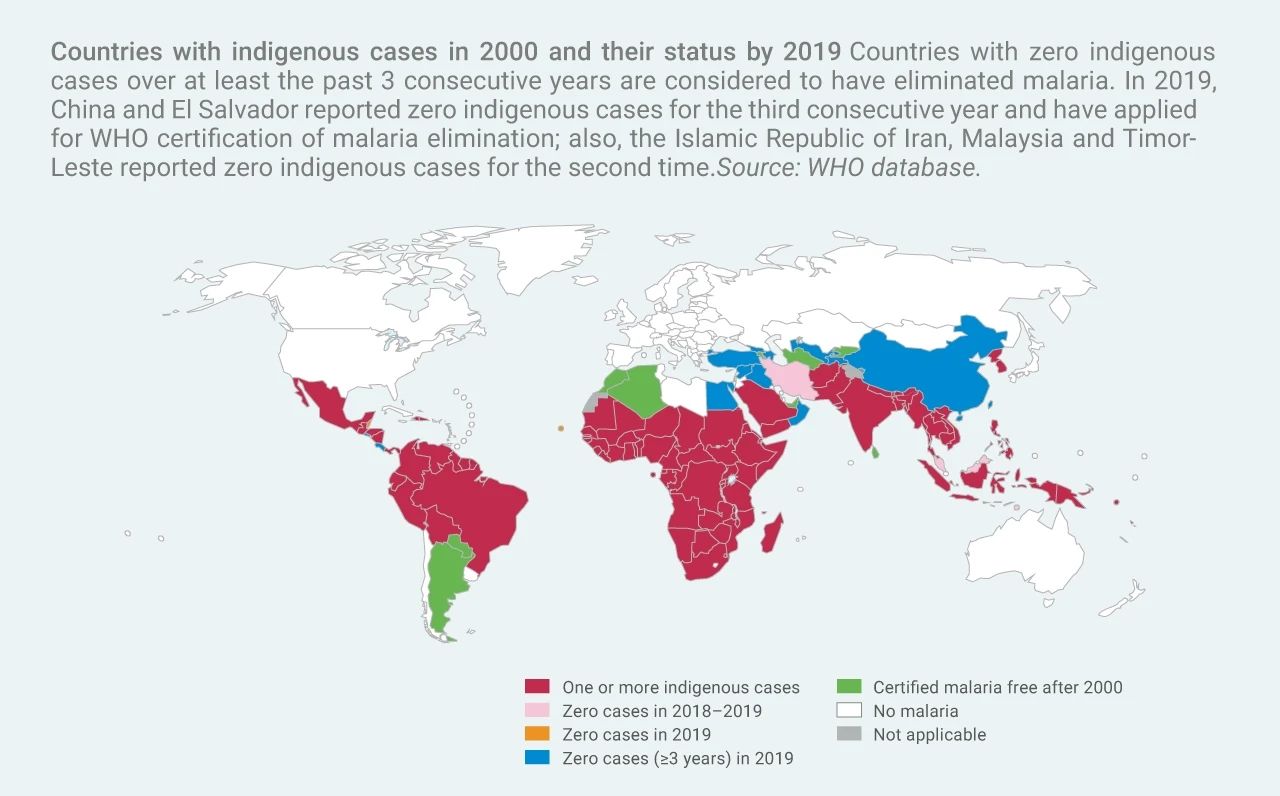
D 1. 2000 ÄęĶвĄĀũĩÄøŧŌŧ°Æä 2019 ÄęĮér (WHO)[1]
¯ÔĪxÉúÃüÖÜÆÚ
¯ŧ˛ĘĮĶɸĐČžČËîĩÄ 5 ˇNŌŅÖĒ¯ÔĪxÖĐĩÄČÎēÎŌģˇNŌũÆđĩÄÔÉúĶÎīŧ˛˛ĄŖēēĐÔ¯ÔĪxĄĸégČÕ¯ÔĪxĄĸČũČÕ¯ÔĪxĄĸÂŅĐίÔĪxĄĸÖZĘĪ¯ÔĪxĄŖÆäÖĐŖŦÔÚÃĀÖŪēÍWÖŪ×îŗŖŌĩÄĘĮégČÕ¯ÔĪxēÍČũČÕ¯ÔĪxŖŦÔÚˇĮÖŪĘĮēĐÔ¯ÔĪxĄŖ
D 2. ŧÄÉúĶÚČËîĩįÔĪxÎīˇN[2]
ģųĶÚžÁŖķwģųŌōŊMĩįÔĪxĪĩŊy°lĶũä
ģųĶÚžÁŖķwģųŌōŊMĩįÔĪxĪĩŊy°lĶũä
¯ÔĪxÉúÃüÖÜÆÚÖ÷ŌĒˇÖéÉëAļÎŖēŧ´°lÉúČËîËŪÖ÷ČĩÄoĐÔˇąÖŗēÍ°lÉúÔÚÎÃĪxÃŊŊéČĩÄĶĐĐÔˇąÖŗĄŖ
oĐÔˇąÖŗëAļÎŖē´ÆĐÔ°´ÎÃĸēŦĶĐ×Ķæß×Ķ (sporozoites) ĩÄÍŲŌē×ĸČëĩŊČËîËŪÖ÷ķwČĄŖÔÚŌģĐĄrČŖŦß@ĐŠ×Ķæß×ĶßMČëŅĒÁ÷˛ĸŅhĩŊ¸ÎÅKŖŦßMļø¸ĐČž¸Îŧ°û (¸Îŧ°ûrÆÚ)ŖŦÔÚ¸Îŧ°ûČŖŦÃŋ×Ķæß×ĶoĐÔÍÖÆĐÎŗÉĩ°ŲÁŅÖŗ×ĶĄŖ¸Îŧ°ûÆÆÁŅŖŦÁŅÖŗ×ĶáˇÅĩŊŅĒŌēÖĐŖŦĮÖČëŧtŧ°û (ŧtŧ°ûÖÜÆÚ)ĄŖ
ÁŅÖŗ×ĶŊvÔįÆÚh î×ĖđBķw (ring)ĄĸÍíÆÚ×ĖđBķw (trophozoite)ŖŦ×´ÎĶĐŊzˇÖÁŅĐÎŗÉÁŅÖŗķw (schizont) ëAļÎĄŖČģēķÁŅÖŗķwÆÆÁŅŖŦĸÁŅÖŗ×ĶáˇÅģØŅĒŌēÖĐŖŦÔÚÄĮĀīËüŋÉŌÔĮÖČëĐÂĩÄŧtŧ°û˛ĸ ĸÅcŧtŧ°ûŅhĄŖ
ĶĐĐÔˇąÖŗëAļÎ: ÉŲĩ×ĖđBķwŪDģ¯éĐÛĐÔēÍ´ÆĐÔÅä×Ķķw (male/female gametocytes)ĄŖŽ´ÆĐÔ°´ÎÃÎüĘŗĘܸĐČžËŪÖ÷ĩÄŅĒŌērŖŦËüūzČĄß@ĐŠÅä×ĶķwŖŦß@ĐŠÅä×ĶķwÔÚÎÃ×ĶķwČßMĐĐĶĐĐÔŅhŖŦĐÎŗÉ×Ķæß×ĶŖŦÍęŗɯÔĪxĩÄÉúÃüÖÜÆÚĄŖ
D 3. ¯ÔĪxÉúÃüÖÜÆÚ[3]
¯ÔĪx¸ĐČžÉúÎīCÖÆ
ŌũÆđ°Y îĐÔ¯ŧ˛ĩÄëAļÎÖ÷ŌĒ°lÉúÔÚ¯ÔĪx¸ĐČžĩÄŧtŧ°ûrÆÚŖŦÔÚ´Ë×ößMŌģ˛ŊĩÄŊéŊBĄŖ
ČëĮÖŖē ÁŅÖŗ×ĶēÍŧtŧ°ûÖŽégĩÄŗõĘŧŊĶĶ|ĘĮëSCĩÄŖŦĩĢéÁËŗÉšĻČëĮÖŖŦÁŅÖŗ×ĶĘšĶÃŧĄĶĩ°°×-ŧĄĮōĩ°°×ß\ĶÖØĐÂļ¨ĪōŖŦĘšÆäíļËÅcŧtŧ°ûĤŊĶĶ|ĄŖÁŅÖŗ×ĶÅcŧtŧ°ûĩÄŗõĘŧĪāģĨ×÷ĶÃÉæŧ°ĩŊÁŅÖŗ×ĶąíÃæĩ°°× 1 (MSP1)ŖŦļøÖØĐÂļ¨ĪōēķĩÄíļ˸ŊÖøÉæŧ°ŧÄÉúĪxĖØļ¨ČëĮÖÅäķw (Čį EBA ēÍ PfRh ŧŌ×åŗÉT) ēÍŧtŧ°ûąíÃæĘÜķwĪāģĨ×÷ĶÃĄŖ
ŗũ´ËÖŽÍâŖŦÁŅÖŗ×ĶēÍŧtŧ°ûĩÄžoÃÜßBŊĶŖŦÅc¯ÔĪxÎĸÁŖĩ°°× AMA1 ēÍŧÄÉúĪxÁâĐÎîi (RON) ĩ°°×ĐÎŗÉĩÄÍēĪÎīŌ˛ĶĐęPĄŖ
D 4. ÁŅÖŗ×ĶČëĮÖ˛ŊķE[5](a) ÁŅÖŗ×ĶÖØĐÂļ¨ĪōĄŖ(b) Åäķw-ĘÜķwĪāģĨ×÷ĶÃŖŦĶÉ EBA ēÍ PfRh ŧŌ×åŗÉTĩÄĩ°°×Ų|Ŋ駥Ŗ(c) PfRh5 ÍēĪÎīÅcĘÜķw basiginĩÄŊYēĪŖģ(d-g) RON ÍēĪÎīŗÁˇeĩŊŧtŧ°ûĤÖĐŖŦÅc AMA-1 ÖąŊĶŊYēĪĐÎŗÉŌÆĶßBŊĶŖŦĸÁŅÖŗ×ĶĄ°ÍÆĄąČëŧtŧ°ûĤŖŦ˛ĸĐÎŗÉŧÄÉúĐÔŌēÅŨĤ (PVM)ĄŖ(h-i) ŧtŧ°ûÖđuģÖ͡BĄŖ
ÍÖÆŖēŌģĩŠŧtŧ°û¸ĐČžąģŊ¨ÁĸŖŦŧÄÉúĪxŅ¸ËŲÍÖÆŖŦÔÚëSēķĩÄ 48 ĐĄrČŖŦÁŅÖŗķwˇÖÁŅŖŦŽaÉúÁŅÖŗ×ĶŖŦß@ĐŠÁŅÖŗ×ĶÔÚ°lĶũrŌŨŗöŖŦ§ÖÂŧtŧ°ûĤÆÆÄēÍŧÄÉúĪxąŦ°lĐÔáˇÅŖŦ˛ĸČëĮÖĐÂĩÄËŪÖ÷ŧ°ûĄŖ
ĶÉĶÚŧÄÉúĪxÔÚŅ¸ËŲÍÖÆß^ŗĖÖĐĐčŌĒŗÖĀmĩÄēËÜÕËáėíēĪŗÉ DNA ēÍ RNAŖŦŌō´ËŖŦÔSļāŋš¯Ë°ĐĪōā×ā¤ÉúÎīēĪŗÉĄŖČį°ĐĪōļūäČ~ËáßÔø (DHFR) ĩÄ PyrimethamineĄĸCycloguanilŖŦ×čāā×ā¤ÉúÎīēĪŗÉĩÄ AtovaquoneŖŦŌÔŧ°Í¨ß^°ĐĪō DHODH í×čāā×ā¤ĩÄÉúÎīēĪŗÉĩÄ DSM265ĄŖ
ÁíÍâŖŦŧÄÉúĪxĻ IđBÎüĘÕĩÄĘÜpÅc Na+ ĩÄĶĐēĻˇeĀÛĶĐęPŖŦļøĩÍĩČÉúÎīČįŧÄÉúĪxĩÄŲ|Ĥ Na+ ŨŗöąÃ (PfATP4)ŖŦŋÉŌÔŧ°rąÃŗöˇeĀÛĩÄ Na+ŖŦÄļøžSŗÖ Na+ ˇBĄŖÁíÍâŖŦ¯ÔĪx PI4K (ČįPfPI4K) ͨß^´ßģ¯Á×ÖŦõŖŧĄ´ŧ (PI) ĩÄÁ×ËáĸĶĪÂĶÎŧÂͨ¡ŖŦÕ{šļāˇNÉúÎīß^ŗĖŖŦ°üĀ¨ÔÚÁŅÖŗ×Ķ°lĶũß^ŗĖÖĐĩÄĐÅĖŪD§ĄĸĤß\ŨēÍ°ûŲ|ˇÖÁŅĄŖ
ÖĩĩÃŌģĖáĩÄĘĮŖŦÄŋĮ°´ķ˛ŋˇÖŋš¯ËÎīļŧĘĮ°ĐĪōŅĒŌēëAļÎŖŦČį Artemisinin ŧ°ÆäŅÜÉúÎīĩČļŧĘĮĶĐЧĩÄŋš¯ËÎīĄŖ
D 5. áĻ¯ÔĪxÉúÃüÖÜÆÚ˛ģÍŦëAļÎĩIJŋˇÖŋš¯Ë[11]
ËŪÖ÷ÃâŌߡ´ĒēÍŧÄÉúĪxÃâŌßĖĶąÜ
ŧÄÉúĪxžßĶĐÆÆÄģōĖĶąÜËŪÖ÷ÃâŌߡ´ĒĩÄÄÜÁĻŖŦŌō´Ëū°lÉúÖØ͸ĐČžĄŖČģļøß@ˇNÆÆÄĩÄCÖÆÄŋĮ°ČÔ˛ģĮåŗūĄŖĶĐŅĐžŋąíÃ÷ŖŦŧÄÉúĪxÄÜÕT§äÍģ îŧ°û (DC) šĻÄÜʧЧŖŦ§ÖÂŽaÉúšĻÄÜʧÕ{ĩÄ T ŧ°ûĄŖß@ĐŠšĻÄÜʧÕ{ĩÄ T ŧ°ûoˇ¨ßwŌÆĩŊÁÜ°ÍŊYÖĐĩÄ B ŧ°û
^ĶōŌÔÍÖú B ŧ°ûÔöÖŗē͡´ĒŖŦÄļøpÉŲÁËŋØÖƯŧ˛¸ĐČžËųąØĐčĩÄŋšķwĩÄŽaÉúĄŖ°l˛ĄŖēÖ÷ŌĒĩÄÖ²ĄCÖÆĘĮ¯ÔĪx¸ĐČžēķŖŦŧtŧ°û°lÉúČÜŅĒŖŦáˇÅŗöŧÄÉúĪxēͯŧ˛ČļžËØ (ąģÕJéĘĮ¯ÔĪxēÍŧÄÉúĪx DNA ĩÄÍēĪÎī)ŖŦßMļøŧ¤ģî TLR9 Ķ|°lŅ×°Yˇ´ĒĄŖ´ËÍâŖŦĘܸĐČžĩÄŧtŧ°ûĩÄĤ×Ķ˛ļøĘʧ×ĐÎÄÜÁĻŖŦß@ˇNÄÜÁĻĩÄĘʧ§ÖÂÃĢŧŅĒšÜ×čČûŖŦŽÖØŌĒÆ÷šŲĘÜĩŊĶ°írŖŦĀÖØrūÎŖŧ°ÉúÃüĄŖ
ŋŊYŖē
žÍÄŋĮ°ļøŅÔŖŦ¯ŧ˛ČÔČģĘĮČĢĮōÖØ´ķĩÄŧ˛˛ĄØúĄŖąMšÜ Artemisinin ŧ°ÆäŅÜÉúÎīĘĮŌģžŋš¯ËÎīŖŦArtemisinin ÂēĪ¯ˇ¨Đ§šû¸üĘĮĘŽˇÖī@ÖøŖŦĩĢÄÍËĐÔŧÄÉúĪxĩÄŗöŦFČÔÅfĘĮÖØŌĒĩÄî}ĄŖŌō´ËŖŦĐčŌĒé_°lĐ°ĐücĩÄĐÂĐÍŋš¯ËŖŦČįģųĶÚŧtŧ°ûČëĮÖĐčŌĒŌģĪĩÁĐ fÕ{ĩÄĩ°°×Ų|/ĩ°°×Ų|ĪāģĨ×÷ĶÃĩČĄŖÁíÍâŖŦÅcŧtŧ°ûąíÃæĩÄ basigin ĪāģĨ×÷ĶÃĩÄēĐÔ¯ÔĪxĩ°°×ÍēĪÎī PfRH5ĄĸPfAMA1 ĩČŌ˛ŋÉ×÷éŌßÃįŅĐžŋĄŖŌĒØĩ×ĪûŗũČËî¯ŧ˛ŖŦÎŌČÔĶĐŌģļΡŌĒ×ߥŖ
| ĪāęPŽaÆˇ |
| Artemisinin ŌģˇN§ĶĐß^ŅõģųFĩÄąļ°ëŨÆČõĨēÍŋš¯ŧ˛ŠĄŖ |
| Chloroquine ͨß^¸É_ŧÄÉúĪxĪûģ¯ŌēÅŨ (food vacuole) íĸËĀŧÄÉúĪxĄŖ |
| Pyrimethamine ßxņĐÔŌÖÖƯŧ˛ļūäČ~ËáßÔøĐØÜÕËáēĪø (DHFR-TS)ŖŦ×čā DNA ēĪŗÉĄŖ |
| Cycloguanil Proguanil ĩÄģîĐÔ´úÖxŽaÎīŖŦ×÷ĶÃĶÚŧtŧ°ûē͸Îŧ°ûÖĐĩįŧ˛ÁŅÖŗķwĄŖ |
| Atovaquone žßĶĐŋÚˇūģîĐÔĩÄßxņĐÔŧÄÉúĪxžÁŖķwŧ°ûÉĢËØ bc1 ÍēĪÎīĩÄŌÖÖÆŠĄŖ |
| Cipargamin ŋėËŲĩÄŋš¯ŠŖŦŌÖÖÆ PfATP4ŖŦĻ¯ÔĪxĩÄËųĶĐŧtŧ°ûČëAļÎļŧĶĐģîĐÔŖŦ˛ĸžßĶĐĸËĀÅä×Ķŧ°ûĩÄģîĐÔŖŦžßĶĐ×čÖš÷˛ĨĩÄÁĻĄŖ |
| MMV390048 ¯ÔĪx PI4K ŌÖÖÆŠĄŖ |
| Antiparasitic Compound library °üēŦ 250+ žßĶĐÃ÷´_ŋšŧÄÉúĪx×÷ĶÃĩÄĐĄˇÖ×Ķģ¯ēĪÎīŖŦŋÉĶÃĶÚĐÂßmĒ°YĩÄé_°lŧ°ĐÂĐÍŋšŧÄÉúĪx°ĐücĩÄčbļ¨ĄŖ |
ĸŋŧÎÄĢI
1. Phillips MA, et al. Malaria. Nat Rev Dis Primers. 2017;3:17050.
2. Escalante AA, et al. Malaria Molecular Epidemiology: An Evolutionary Genetics Perspective. Microbiol Spectr. 2019;7(4):10.1128/microbiolspec. AME-0010-2019.
3. De Koning-Ward TF, et al. Plasmodium species: master renovators of their host cells. Nat Rev Microbiol. 2016;14(8):494-507.
4. Zheng Z, et al. Advances in Molecular Diagnosis of Malaria. Adv Clin Chem. 2017;80:155-192.
5. Cowman AF, et al. Malaria: Biology and Disease. Cell. 2016;167(3):610-624.
6. Mohandas N, et al. Malaria and human red blood cells. Med Microbiol Immunol. 2012;201(4):593-598.
7. Liang X, et al. Discovery of 6'-chloro-N-methyl-5'-(phenylsulfonamido)-[3,3'-bipyridine]-5-carboxamide (CHMFL-PI4K-127) as a novel Plasmodium falciparum PI(4)K inhibitor with potent antimalarial activity against both blood and liver stages of Plasmodium. Eur J Med Chem. 2020;188:112012.
8. Hisaeda H, et al. Malaria: immune evasion by parasites. Int J Biochem Cell Biol. 2005;37(4):700-706.
9. Osii RS, et al. The Impact of Malaria Parasites on Dendritic Cell-T Cell Interaction. Front Immunol. 2020;11:1597. Published 2020 Jul 24.
10. Yang J, et al. Advances in the research on the targets of anti-malaria actions of artemisinin. Pharmacol Ther. 2020;216:107697.
11. Delves M, et al. The activities of current antimalarial drugs on the life cycle stages of Plasmodium: a comparative study with human and rodent parasites. PLoS Med. 2012;9(2):e1001169.

- ŧ°ûčFËĀÍöŗŖĶÃŌÖÖÆŠĄĸÕT§Šŧ°zyˇŊˇ¨ēÍĒĶð¸Āũ
- AcetylcysteineŖ¨NACŖŠĩÄ×÷ĶÃCÖÆŧ°ÔÚŋšŅõģ¯ĩČŅĐžŋÖĐĩÄĒĶÃ
- Mitomycin CĩÄ×÷ĶÃCÖÆŧ°ÔÚÄ[Áöŧ°ûĪĩēÍĶÎīÄŖĐÍÖĐĩÄļāöž°ĒĶÃ
- CFDA-SEĩÄ×÷ĶÃCÖÆŧ°ÔÚŧ°ûÔöÖŗēÍķwČĘžÛÖĐĩÄĒĶÃ
- ŌÖÖÆŠA83-01ĩÄ×÷ĶÃCÖÆŧ°ÔÚ¸Éŧ°ûÅcîÆ÷šŲÅāđBÖĐĩÄĒĶÃ
- 2-Deoxy-D-glucoseŖ¨2-DGŖŠĩÄ×÷ĶÃCÖÆŧ°ÔÚĖĮŊÍŊâŌÖÖÆŅĐžŋÖĐĩÄ×÷ĶÃ
- â}ëx×ĶÉšâĖŊáCalbryte 520.AMÔÚĶBąOyÖĐĩÄēËĐÄŨÅcĒĶÃŊâÎö
- NRF2ēÍAKTŧ¤ģîģōFBP1ȹʧÔÚ¸Îŧ°ûËĨĀĪē͸ΰŠßMÕšÖĐĩÄ×÷ĶÃ
- ×ĸÔūTŖŦAbMoleŧ°ûō4´ķŊđÔŠČÎŌģŋîÃâŲMËÍ
- MCEÁųÔÂÕšūîA¸æŖēČËš¤ÖĮÄÜ´ķū&¸Éŧ°ûŅĐžŋū×h
- MCE HOLOĪĩÁĐžųĪāÃâŌߡÖÎöÔŠēĐÉĪĐÂ5ÕÛLõrr
- MCEÖą˛ĨîA¸æŖēPROTACŅĐžŋĐÂßMÕšēÍÔOĶĐÂËŧ¡
- MCEļËÎįëpÖظŖĀûŖēĘÖËŲPKŲēÍáéTļāëÄŽaÆˇMŲļYÆˇ
- MCEÖĐøŗõĪÄ´ŲäNģîĶŖēČžÁĪŽaÆˇâ ģŨ55ÕÛŋņg
- MCEÅcÄãĪāŧsĩÚÁųÃøëHŧ°ûÅcģųŌōÖίÖĐøˇåū
- Î÷°˛°ŲÎ618ÄęÖĐ´ķ´Ų,ŲIÉšâČžÁĪË͸ßͨÁŋŖĶÃÎĸŋ×°å


Copyright(C) 1998-2025 ÉúÎīÆ÷˛ÄžW ëÔŖē021-64166852;13621656896 E-mailŖēinfo@bio-equip.com