Ц¬·ҫЛбFAФЪуwғИ…ўЕcОьКХкPжIХ{ҝШҷCЦЖөДЦШТӘЧчУГ

Ц¬·ҫЛбЈЁFAЈ©ФЪуwғИҫЯУРЦШТӘ№ҰДЬЈ¬ҝЙЧчһйДЬБҝҒнФҙЈ¬Іў…ўЕcЙъОпДӨәПіЙәНДЬБҝҙжғҰЎЈИ»¶шЈ¬Ц¬·ҫЛбИзәОҝзЯ^јҡ°ыДӨЯMИлјҡ°ығИЈ¬ДҝЗ°ИФІ»М«ЗеіюЎЈЕcЖПМСМЗәН°ұ»щЛбІ»Н¬Ј¬Ц¬·ҫЛбҫЯУРКиЛ®РФЈ¬Я@К№өГЖдЯ\„УлyТФЧ·ЫҷЎЈУРИЛХJһйЈ¬Ц¬·ҫЛбКЗҪӣЯ^ұ»„У”UЙўҙ©Я^јҡ°ыДӨЈ¬ө«ФҪҒнФҪ¶аөДЧC“юұнГчЈ¬Ц¬·ҫЛбКЗФЪө°°ЧөД…ўЕcПВНкіЙҙъЦxҪMҝ—өДҝзДӨЮDЯ\ЎЈ
ФЪТСЦӘөДЦ¬·ҫЛбЮDЯ\ө°°ЧЦРЈ¬CD36КЗСРҫҝПаҢҰЭ^¶аөДТ»ӮҖЎЈЛьЦчТӘ¶ЁО»УЪјҡ°ыДӨЙПөДcaveolae°јПЭҪYҳӢЈ¬ЗТЦ¬·ҫЛбОьКХ»оРФТІТАЩҮcaveolaeҪYҳӢЎЈРЎКуЦ¬·ҫәНјЎИвҪMҝ—ЦРҙујs50%өДЦ¬·ҫЛбОьКХКЗУЙCD36Ш•«IөДЎЈCD36УРТ»ӮҖЦ¬·ҫЛбҪYәПО»ьcЈ¬ө«ҫЯуwИзәОЮDЯ\Ц¬·ҫЛбЈ¬¬FФЪЯҖКЗӮҖЦiЎЈ
ҸНө©ҙуҢWҙъЦxЕcХыәПЙъОпҢWСРҫҝФәөДЪwН¬ҪрХnо}ҪMЦ®З°°l¬FЈ¬ғЙӮҖЧШҷ°хЈ»щЮDТЖГёDHHC4әНDHHC5·Ц„eФЪёЯ –»щуwәНјҡ°ыЩ|ДӨЙПҢҰCD36ЯMРРЧШҷ°хЈ»ҜРЮп—Ј¬ҸД¶шҫSіЦCD36өДЩ|ДӨ¶ЁО»ІўҙЩЯMЖдЦ¬·ҫЛбОьКХөД»оРФЎЈ
ЧоҪьЈ¬ЪwН¬ҪрХnо}ҪMЕcДПҫ©ҙуҢWДЈКҪ„УОпСРҫҝЦРРДөДкҗҺӣХnо}ҪMәПЧчЈ¬ЯMТ»ІҪСРҫҝБЛCD36ҙЩЯMЦ¬·ҫЛбОьКХөД·ҪКҪЎЈЛыӮғ°l¬FЈ¬Ц¬·ҫЛбҝЙНЁЯ^CD36ҪйҢ§өДғИНМЧчУГЮDЯ\өҪјҡ°ығИЈ¬¶шCD36өД„У‘BЧШҷ°хЈ»ҜФЪЯ@Т»Я^іМЦР°l“]кPжIЧчУГЎЈЯ@н—іЙ№ы°lұнФЪЎ¶Nature CommunicationsЎ·лsЦҫЙПЎЈ

CD36ҪйҢ§өДғИНМЧчУГҢўЦ¬·ҫЛбЮDЯ\өҪјҡ°ы
һйБЛСРҫҝЦ¬·ҫЛбОьКХЖЪйgCD36өДҒҶјҡ°ы¶ЁО»Чғ»ҜЈ¬СРҫҝИЛҶTУГBSAЕјВ“өДУНЛбМҺАн3T3-L1Ц¬·ҫјҡ°ыЈ¬Іўй_Х№ғИФҙРФCD36өДГвТЯИҫЙ«ЎЈЛыӮғ°l¬FЈ¬ҸДУНЛбМҺАнәу30·ЦзҠй_КјЈ¬CD36ұ»ғИ»ҜІўФЪјҡ°ығИп@Кҫіцьc оРЕМ–ЎЈөҪ1РЎ•rЈ¬CD36й_Кј°ь№ьЦ¬өОЈ¬ІўФЪ2РЎ•rәН4РЎ•rЧғөГёьГчп@ЎЈ
УЙУЪCD36ЦчТӘ¶ЁО»ФЪcaveolae°јПЭҪYҳӢЙПЈ¬№КСРҫҝИЛҶTТІ·ЦОцБЛcaveolaeҪYҳӢө°°ЧCaveolin1ЈЁCAV1Ј©өД„У‘B¶ЁО»ЎЈЛыӮғ°l¬FCAV1ЕcCD36№ІН¬ЯwТЖЎЈҝј‘]өҪCD36өДЦ¬·ҫЛбҪYәПО»ьcО»УЪЩ|ДӨНвӮИЈ¬¶шCAV1О»УЪғИӮИЈ¬ЛыӮғІВңyCD36әНCAV1ФЪғИ»Ҝ•rПа»ҘТАЩҮЈ¬ЖдЦРCD36ЧчһйКЬуwҪУКЬЦ¬·ҫЛбөДРЕМ–Ј¬¶шCAV1МṩҪYҳӢЙПөДЦ§іЦЎЈҙ_ҢҚКЗЯ@ҳУЈ¬®”Ц¬·ҫјҡ°ыЦРөДИОТ»ӮҖұ»ЗГіэ•rЈ¬БнТ»ӮҖФЪУНЛбМҺАнәуТІІ»ФЩғИ»ҜЈ¬Я@ұнГчЛьӮғөДғИ»ҜФҙУЪCD36ҪйҢ§өДcaveolaeғИНМЧчУГЎЈ
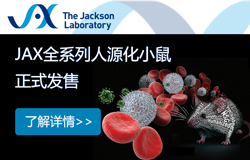
Ц®әуЈ¬ЛыӮғЯx“сPacFAЧчһйЦ¬·ҫЛбоҗЛЖОпҒнЧ·ЫҷЦ¬·ҫЛбФЪјҡ°ығИөДЯ\„УЎЈИзҲD1ЛщКҫЈ¬PacFAКЗТ»·NҝЙТФ№в»о»ҜөДЦ¬·ҫЛбоҗЛЖОпЈ¬ҝЙ…ўЕcьc“ф·ҙ‘ӘЎЈЛыӮғУГPacFAМҺАнЦ¬·ҫјҡ°ы20·ЦзҠЈ¬И»әуАыУГN3-Alexa Fluor 488й_Х№ьc“ф·ҙ‘ӘТФҳЛУӣPacFAЈ¬ІўҢҰCD36ЯMРРГвТЯИҫЙ«ЎЈНЁЯ^Я@·N·ҪКҪЈ¬ЛыӮғУ^ІмөҪБЛPacFAәНCD36өДЗеОъ¶ЁО»ЎЈФЪ¶ЁБҝ•rЈ¬ҙујs56%өДPacFAЕcјҡ°ығИІҝөДCD36ҪYәПЎЈ
һйБЛЯMТ»ІҪҙ_ХJCD36НЁЯ^ғИНМЧчУГҙЩЯMЦ¬·ҫЛбОьКХЈ¬СРҫҝИЛҶTФҮҲDЗГіэCAV1Ј¬ҝҙҝҙҢҰғИНМЧчУГөДЧи”аКЗ·с•юЖЖүДЦ¬·ҫЛбОьКХЎЈЛыӮғФЪCav1−/−РЎКуЈЁЩҗҳIЙъОпМṩЈ©Ўў Cd36−/−РЎКуТФј° Cav1−/−;Cd36−/−РЎКуөДЦ¬·ҫјҡ°ыЦРй_Х№3H-УНЛбОьКХҢҚтһЎЈЕcТ°ЙъРНЦ¬·ҫјҡ°ыПаұИЈ¬Cav1−/−Ц¬·ҫјҡ°ыЦРөД3H-УНЛбОьКХҪөөН40%ЎЈН¬•rЈ¬лpЦШЗГіэІўӣ]УРЯMТ»ІҪҪөөН3H-УНЛбөДОьКХЈ¬ұнГчcaveolarғИНМКЗCD36ТАЩҮөДЦ¬·ҫЛбОьКХЛщұШРиөДЎЈ
ҲD1. CD36ҪйҢ§өДcaveolarғИНМҢўЦ¬·ҫЛбЮDЯ\өҪјҡ°ы
CD36өД„У‘BЧШҷ°хЈ»ҜКЗЦ¬·ҫЛбОьКХЛщұШРиөД
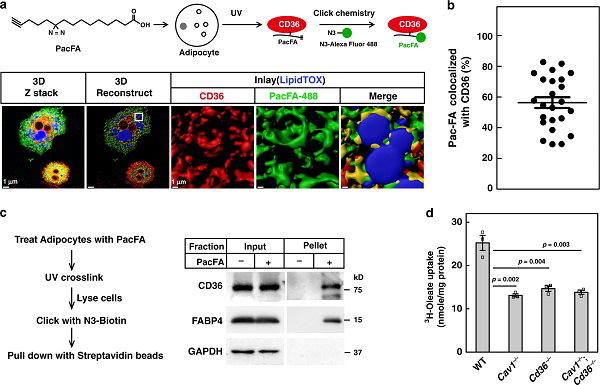
Ц®әуЈ¬СРҫҝИЛҶTПлБЛҪвCD36ИзәОҸДЩ|ДӨЙПҪвлxЎЈУЙУЪCD36өДЩ|ДӨ¶ЁО»РиТӘЧШҷ°хЈ»ҜЈ¬ТтҙЛЛыӮғПлҝҙҝҙCD36ФЪғИ»ҜЦ®З°КЗ·с•юИҘЧШҷ°хЈ»ҜЎЈФЪУНЛбМҺАнәу5·ЦзҠғИЈ¬CD36өДЧШҷ°хЈ»ҜЛ®ЖҪй_КјҪөөНЈ¬ІўФЪ30-60·ЦзҠЧуУТЯ_өҪЧоөНьcЎЈDHHC5өДЧШҷ°хЈ»ҜТІУРоҗЛЖөДУ^ІмҪY№ыЎЈҸДЕарB»щЦРИҘіэУНЛб•rЈ¬CD36әНDHHC5ұ»ЦШРВЧШҷ°хЈ»ҜЈ¬Я@ұнГчCD36әНDHHC5ФЪЦ¬·ҫЛбХTҢ§өДғИНМЯ^іМЦРҪӣҡvБЛЧШҷ°хЈ»ҜөД„У‘BЧғ»ҜЎЈ
ҪУЦшЛыӮғҪвОцБЛCD36ғИНМөДРЕМ–НЁВ·ЎЈКЧПИиb¶ЁБЛCD36өДИҘЧШҷ°хЈ»ҜГёЈ¬ДҝЗ°ЦчТӘУРОе·NЈ¬°ьАЁAPT1ЎўAPT2әНABHD17A–CЎЈНЁЯ^PalmBТЦЦЖөИҢҚтһЈ¬ЛыӮғ°l¬FAPT1КЗCD36өДИҘЧШҷ°хЈ»ҜГёЎЈИ»әуЈ¬ЛыӮғЗГіэБЛ3T3-L1Ц¬·ҫјҡ°ыЦРөДAPT1»тІЙУГAPT1ТЦЦЖ„©МҺАнЈ¬°l¬FAPT1өДЗГіэ»тТЦЦЖҫщҝЙЧи”аУНЛбХTҢ§өДCD36ғИ»ҜәНИҘЧШҷ°хЈ»ҜЈЁҲD2Ј©ЎЈ
ҲD2. CD36өДИҘЧШҷ°хЈ»ҜКЗЦ¬·ҫЛбғИНМЛщұШРиөД
ДЗГҙКЗКІГҙҢ§ЦВБЛCD36өДИҘЧШҷ°хЈ»ҜЎЈСРҫҝИЛҶTІВПлЈ¬DHHC5ҝЙДЬұ»ңз»оЈ¬ТтһйDHHCөДЧФЙнЧШҷ°хЈ»ҜКЗҙЯ»ҜөЧОпЧШҷ°хЈ»ҜөДөЪТ»ІҪЎЈәуАmөДСРҫҝ°l¬FЈ¬ФЪУНЛбМҺАн5·ЦзҠәуЈ¬DHHC5ЙПTyr91О»ьcөДБЧЛб»Ҝп@ЦшФцјУЈ¬¶ш30·ЦзҠәуҪөЦБ»щөЧЛ®ЖҪЎЈЛыӮғЯҖЧCҢҚЈ¬DHHC5өД„У‘BК§»оКЗCD36ғИ»ҜЛщұШРиөДЎЈ
УЪКЗЈ¬ПВТ»ӮҖДҝҳЛКЗиb¶ЁҢўDHHC5БЧЛб»ҜөДјӨГёЎЈЛыӮғЗГіэБЛ3T3-L1Ц¬·ҫјҡ°ыЦРШS¶ИПаҢҰЭ^ёЯөДЛДӮҖSrcјТЧејӨГёЈЁSFKЈ©Ј¬°l¬FLYNөДЗГіэҝЙТЦЦЖУНЛбХTҢ§өДDHHC5БЧЛб»ҜЎЈАыУГғЙӮҖshRNAЗГіэLYNәуЈ¬УНЛбХTҢ§өДCD36ғИ»ҜТФј°CD36әНDHHC5өДИҘЧШҷ°хЈ»Ҝұ»Чи”аЈ¬ЧCҢҚLYNҝЙФЪTyr91О»ьcҢўDHHC5БЧЛб»ҜЎЈ
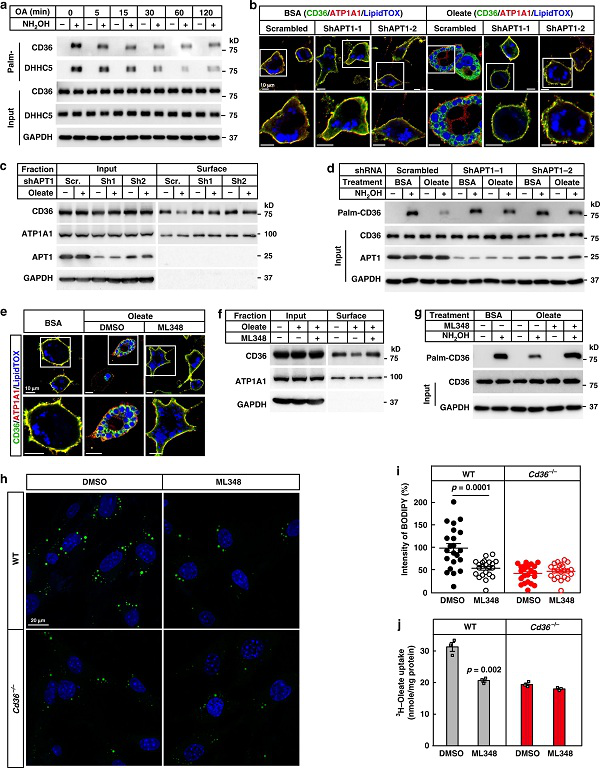
І»Я^Ј¬CD36өДИҘЧШҷ°хЈ»ҜИзәОҶў„УғИНМЧчУГДШЈҝСРҫҝИЛҶTКЧПИСРҫҝБЛdynaminЎўVAVәНJNKөДХРДјЈ¬Я@Р©ө°°ЧФшұ»ҲуөА…ўЕcБЛCD36өДғИНМЧчУГЎЈАыУГёчӮҖө°°ЧөДТЦЦЖ„©МҺАнәуЈ¬ЛыӮғ°l¬FЛщУРТЦЦЖ„©ҫщДЬЧи”аУНЛбХTҢ§өДCD36ғИНМЧчУГЎЈН¬•rЈ¬УНЛбМҺАнҢ§ЦВVAVәНJNKөДБЧЛб»ҜФцјУЈ¬ұнГчCD36өДИҘЧШҷ°хЈ»ҜРиТӘХРДјЯ@Р©ө°°ЧЎЈ
НЁЯ^ЯMТ»ІҪөДСРҫҝЈ¬ЛыӮғ°l¬FАТ°ұЛбјӨГёSYKФЪУНЛбМҺАнәуұ»јӨ»оЈ¬ІўХРДјөҪЩ|ДӨЙПЎЈSYKТЦЦЖ„©ЈЁpiceatannolЈ©өДМҺАнФЪәЬҙуіМ¶ИЙППыіэБЛУНЛбХTҢ§өДVAVәНJNKБЧЛб»ҜТФј°CD36өДғИ»ҜЈ¬ө«ІўІ»У°н‘CD36ИҘЧШҷ°хЈ»ҜЈ¬ұнГчSYKјӨ»оКЗCD36ИҘЧШҷ°хЈ»ҜөДПВУОКВјюЎЈSYKҢўVAVәНJNKБЧЛб»ҜЈ¬ҸД¶шҶў„УCD36ҪйҢ§өДғИНМЧчУГЎЈ
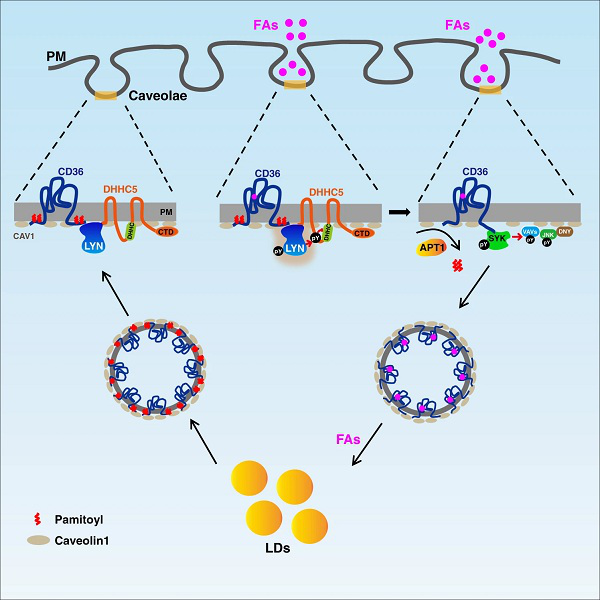
ғИНМЧчУГөДЧи”аҝЙТЦЦЖCD36өДЙъАн№ҰДЬ
ЧоәуЈ¬СРҫҝИЛҶTФu№АБЛCD36ҪйҢ§өДЦ¬·ҫЛбғИНМУРәОЙъАнТвБxЎЈУЙУЪЦ¬·ҫЛбОьКХөДЦчТӘ№ҰДЬКЗҢўЦ¬·ҫЛбғҰҙжФЪЦ¬·ҫјҡ°ыөДЦ¬өОЦРЈ¬ТтҙЛЛыӮғМҪУ‘БЛғИНМЧчУГөДЧи”аҢҰЦ¬өОЙъйLУРәОУ°н‘ЎЈЕcТ°ЙъРНЦ¬·ҫјҡ°ыІ»Н¬Ј¬Cd36−/−Ц¬·ҫјҡ°ыҪӣЯ^УНөОМҺАнәуҺЧәхӣ]УРГчп@өДЦ¬өОЙъйLЈ¬ұнГчғИНМЧчУГөДЧи”аҝЙТФПыіэCD36ТАЩҮРФөДЦ¬өОЙъйLЎЈҙЛНвЈ¬НЁЯ^°РПтLYN»тSYKҒнЧи”ағИНМЧчУГЈ¬ТІДЬүтТЦЦЖпӢКіХTҢ§·КЕЦРЎКуөДуwЦШФцјУЎЈ
ЧоәуЈ¬СРҫҝИЛҶTФu№АБЛCD36ҪйҢ§өДЦ¬·ҫЛбғИНМУРәОЙъАнТвБxЎЈУЙУЪЦ¬·ҫЛбОьКХөДЦчТӘ№ҰДЬКЗҢўЦ¬·ҫЛбғҰҙжФЪЦ¬·ҫјҡ°ыөДЦ¬өОЦРЈ¬ТтҙЛЛыӮғМҪУ‘БЛғИНМЧчУГөДЧи”аҢҰЦ¬өОЙъйLУРәОУ°н‘ЎЈЕcТ°ЙъРНЦ¬·ҫјҡ°ыІ»Н¬Ј¬Cd36−/−Ц¬·ҫјҡ°ыҪӣЯ^УНөОМҺАнәуҺЧәхӣ]УРГчп@өДЦ¬өОЙъйLЈ¬ұнГчғИНМЧчУГөДЧи”аҝЙТФПыіэCD36ТАЩҮРФөДЦ¬өОЙъйLЎЈҙЛНвЈ¬НЁЯ^°РПтLYN»тSYKҒнЧи”ағИНМЧчУГЈ¬ТІДЬүтТЦЦЖпӢКіХTҢ§·КЕЦРЎКуөДуwЦШФцјУЎЈ
ҲD3. CD36ҪйҢ§өДcaveolarғИНМҙЩЯMЦ¬·ҫЛбОьКХөД№ӨЧчДЈРН
ФӯОДҷzЛчЈә
Hao, J., Wang, J., Guo, H. et al. CD36 facilitates fatty acid uptake by dynamic palmitoylation-regulated endocytosis. Nat Commun 11, 4765 (2020). https://doi.org/10.1038/s41467-020-18565-8.
ЩҗҳIOriCellөЪИэҢГјҡ°ыЕарBҲDЖ¬ҙуЩҗй_КјАІ

ЖҜББөДјҡ°ыҝӮКЗПаЛЖөДЈ¬¶ш№·Һ§өДјҡ°ыёчУРёчөДҫ«ІКЎЈХэ·кТ»ДкТ»¶ИөДЩҗҳIOriCellөЪИэҢГјҡ°ыЕарBҲDЖ¬ҙуЩҗЈ¬ОТӮғХчјҜБЛИәАпРЎ»п°йӮғөДҪЁЧhЈ¬Я@ҙОөДұИЩҗІ»ғHұИХlрBөГјҡ°ыёьЖҜББЈ¬ЯҖұИХlрBјҡ°ыёь‘KЈ¬ЧҢҙујТҸДіЙ№ҰәНК§”ЎХэ·ҙлpГжҝӮҪYјҡ°ыЕарBөДҪӣтһҪМУ–ЎЈҝм’ЯГиПВ·Ҫ¶юҫSҙa…ўјУ“OriCellөЪИэҢГјҡ°ыЕарBҲDЖ¬ҙуЩҗ”°ЙЈ¬ФЪ·ЦПнҪӣтһФмёЈҸVҙуҝЖСРНфөДН¬•rЈ¬ЯҖҝЙТФЪAөГРД„УҙуӘ„ЎЈ


- ҸДСЧ°YөҪјІІЎЦОҜҹөДкPжIҪЗЙ«°Чјҡ°ыҪйЛШIL-6әҶҪйј°ПакPРЎКуДЈРНөД‘ӘУГ
- IL-23јҡ°ыТтЧУХTҢ§ДЈРНФЪЦъБҰгyРјІЎСРҫҝЦРөД‘ӘУГ
- ЧVПөКҫЫҷПөБРөЪ¶юЖЪЦ®ҶОЦШҪMГёҪйҢ§өД¶аЙ«ҲуёжПөҪy·Ҫ·Ё
- Bјҡ°ытҢ„УөДEAEДЈРНФЪЦъБҰ¶а°lРФУІ»ҜСРҫҝН»ЖЖЦРөД‘ӘУГ
- M-NSGЦШ¶ИГвТЯИұПЭРЎКуФЪҪТКҫјҡ°ыҙъЦxЦШҫҺіМҷCЦЖЦРөД‘ӘУГ
- РЎКуҙуҢWҶ–Ц®Cre-loxПөҪyөДіЈТҠҶ–о}ЕcҪвҙр…RҝӮ
- іЈИҫЙ«уwл[РФТ•ҫWДӨЙ«ЛШЧғРФЈЁarRPЈ©ПакPРЎКуДЈРНRho KOРЎКуөДҪйҪB
- ҸДҪЁДЈөҪЦОУъЈәҫҖБЈуwТЖЦІіЙ№ҰДжЮDРЎДXНЛРРРФјІІЎДЈРНРЎКу°Y оөДСРҫҝ
- ДПДЈЦұІҘЈәТ»ЧчҪвЧxЕБҪрЙӯІЎЦОҜҹРВ°РьcFAM171A2
- ДПДЈЙъОпЕcGTPСР°lЦРРДХэКҪәһКрGTPәПЧчНЖҸVУӢ„қ
- ГчСёЙъОпЧФЦчСР°lTIGITИЛФҙ»ҜРЎКуДЈРНЦШ°хҒнТu
- ДПДЈЙъОпҢў”y¶аӮҖ„УОпДЈРНЧоРВСРҫҝіЙ№ыББПаAACRДк•ю
- ДПДЈЙъОпСыДъПајsИA–|өЪ17ҢГҢҚтһ„УОпҝЖҢWҢWРgҪ»Бч•ю
- ДПДЈЙъОпҙу/РЎКуҢҚтһІЩЧчҫҖПВЕаУ–°аөЪ¶юЭҶҲуГыНЁЦӘ
- ДПДЈЙъОпСыДъПајsөЪ¶юҢГЧФГвЛҺОпј°Ҝҹ·ЁХ“үҜ
- ёЯЖ·Щ|ҢҚтһЧҰуёЙПРВЈЎұұҫ©әЈНюЕНКҜ«@ңКЯMҝЪ·ЗЦЮЧҰуё