
Plant Physiol:DAP-seq助力揭示番茄果實(shí)成熟調(diào)控新機(jī)制
研究背景
番茄是全球廣泛種植的作物,其肉質(zhì)果實(shí)富含類胡蘿卜素、維生素和酚類化合物,具有重要的經(jīng)濟(jì)價(jià)值。成熟是一個(gè)復(fù)雜且受到嚴(yán)格調(diào)控的發(fā)育過程,會(huì)顯著改變果實(shí)的顏色、質(zhì)地、風(fēng)味和香氣。這一過程由植物激素和遺傳調(diào)控因子之間的復(fù)雜相互作用驅(qū)動(dòng)。C2H2 型鋅指轉(zhuǎn)錄因子在植物生長(zhǎng)、發(fā)育和抗逆性中具有重要作用,但其在果實(shí)成熟中的具體功能仍缺乏研究。
文獻(xiàn)解讀
2025年1月,中國農(nóng)業(yè)大學(xué)園藝學(xué)院郭仰東/張娜團(tuán)隊(duì)在Plant Physiology(IF=6.5)發(fā)表了題為“Protein phosphatase PP2C2 dephosphorylates transcription factor ZAT5 and modulates tomato fruit ripening”的研究論文,該研究揭示了SlPP2C2-SlZAT5模塊介導(dǎo)的蛋白去磷酸化修飾在調(diào)節(jié)呼吸躍變型果實(shí)成熟中的作用,本文借助DAP-seq技術(shù),在全基因組范圍內(nèi)系統(tǒng)鑒定了SlZAT5的結(jié)合位點(diǎn),篩選出其直接調(diào)控的靶基因,解析了SlZAT5的轉(zhuǎn)錄調(diào)控機(jī)制,為改良番茄果實(shí)品質(zhì)提供了理論支撐。

技術(shù)路線
研究結(jié)果
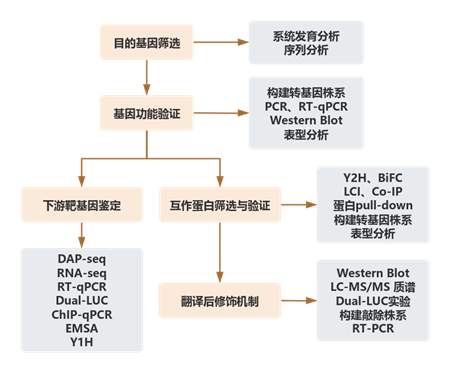
先前研究發(fā)現(xiàn)AdZAT5是獼猴桃果實(shí)軟化的潛在調(diào)節(jié)因子。本研究通過系統(tǒng)發(fā)育分析,在番茄中找到其同源基因SlZAT5,該基因含兩個(gè)保守C2H2結(jié)構(gòu)域和EAR基序。RT-qPCR結(jié)果表明SlZAT5在番茄各組織均有表達(dá),果實(shí)未成熟綠色階段表達(dá)較高、成熟時(shí)下降。SlZAT5突變體開花至破口期縮短至約43天,乙烯峰值提前、硬度下降、類胡蘿卜素積累高;SlZAT5過表達(dá)株系則延長(zhǎng)至約47天,乙烯峰值延遲、硬度增加、類胡蘿卜素積累低,表明SlZAT5負(fù)調(diào)控番茄果實(shí)成熟。
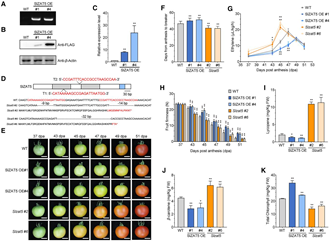
圖1. SlZAT5負(fù)調(diào)控番茄果實(shí)成熟。
為了確定SlZAT5在果實(shí)成熟過程中的直接轉(zhuǎn)錄靶標(biāo),作者通過DAP-seq分析,在7754個(gè)基因中檢測(cè)到SlZAT5結(jié)合峰(高可信度區(qū)域)。結(jié)合RNA-seq數(shù)據(jù),篩選出1511個(gè)受SlZAT5調(diào)控的成熟相關(guān)基因,其中包括乙烯合成基因SlACS4、細(xì)胞壁降解基因SlPL8和轉(zhuǎn)錄因子基因SlGRAS38等關(guān)鍵基因,這些基因的啟動(dòng)子區(qū)域均有SlZAT5結(jié)合位點(diǎn)富集。進(jìn)一步通過RT-qPCR、Y1H、Dual-Luc、ChIP-qPCR和EMSA多種實(shí)驗(yàn)證實(shí),SlZAT5可直接結(jié)合上述基因啟動(dòng)子并抑制其表達(dá)。研究表明SlZAT5通過結(jié)合關(guān)鍵基因啟動(dòng)子調(diào)控果實(shí)成熟過程。
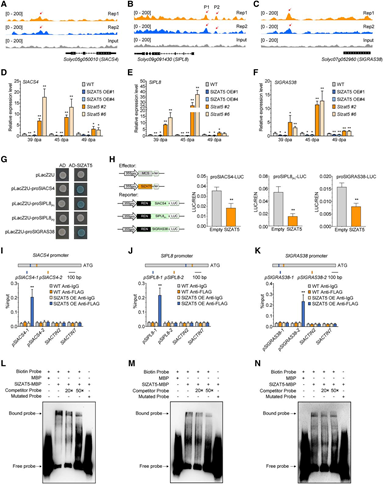
圖2. SlZAT5直接抑制與成熟相關(guān)的基因表達(dá)。
為進(jìn)一步探究SlZAT5在果實(shí)成熟中的作用,作者通過Y2H文庫篩選,鑒定出其互作蛋白SlPP2C2。同時(shí)通過BiFC、LCI、蛋白pull-down、Co-IP、蛋白亞細(xì)胞共定位多種實(shí)驗(yàn),證實(shí)了二者的相互作用,且發(fā)生在細(xì)胞核內(nèi)。通過酵母轉(zhuǎn)錄激活實(shí)驗(yàn)及原生質(zhì)體報(bào)告系統(tǒng)發(fā)現(xiàn),SlZAT5全長(zhǎng)及含EAR基序的C端截短片段均無轉(zhuǎn)錄激活活性,但可抑制熒光素酶活性,表明其作為轉(zhuǎn)錄抑制因子發(fā)揮功能。以上結(jié)果明確了SlZAT5與SlPP2C2的互作關(guān)系及SlZAT5的轉(zhuǎn)錄抑制特性。
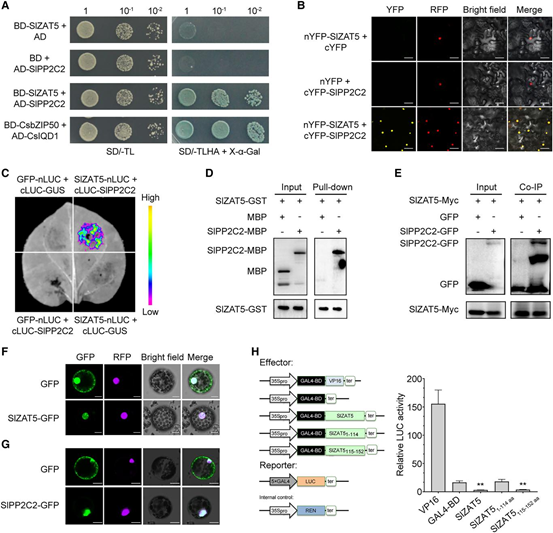
圖3. 轉(zhuǎn)錄抑制因子SlZAT5與SlPP2C2存在物理相互作用。
通過RT-qPCR檢測(cè)SlPP2C2在不同組織和果實(shí)成熟階段的表達(dá),發(fā)現(xiàn)其在果實(shí)未成熟綠色(IMG)和成熟綠色(MG)階段高表達(dá),隨成熟進(jìn)程下降。研究構(gòu)建了Slzat5、Slpp2c2和Slzat5-Slpp2c2突變體:Slpp2c2突變體果實(shí)成熟加速,乙烯水平、類胡蘿卜素含量升高,硬度和葉綠素含量降低。Slzat5-Slpp2c2雙突變體,經(jīng)水處理果實(shí)成熟更快;而經(jīng)乙烯、ACC和1-MCP處理,突變體與野生型的表型無明顯差異。結(jié)果表明SlZAT5和SlPP2C2通過調(diào)控乙烯合成控制番茄果實(shí)成熟。
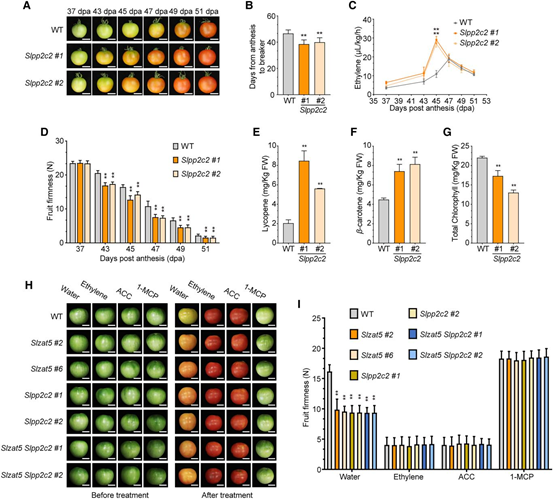
圖4. SlZAT5和SlPP2C2通過調(diào)節(jié)乙烯反應(yīng)來調(diào)控番茄果實(shí)的成熟。
為探究SlZAT5是否受SlPP2C2翻譯后修飾,通過WB檢測(cè)發(fā)現(xiàn),SlZAT5過表達(dá)樣本中存在SlZAT5蛋白及其磷酸化形式,經(jīng)磷酸酶處理后磷酸化信號(hào)消失;SlPP2C2-GFP與SlZAT5-MYC共表達(dá)時(shí),未檢測(cè)到磷酸化條帶。進(jìn)一步通過LC-MS/MS分析,確定了Ser-65為磷酸化位點(diǎn)。雙熒光素酶報(bào)告實(shí)驗(yàn)表明,SlZAT5及去磷酸化突變體S65A可抑制下游基因活性,而磷酸化突變體S65D無此作用。此外,敲除SlPP2C2顯著上調(diào)目標(biāo)基因轉(zhuǎn)錄水平。以上結(jié)果表明,SlPP2C2通過去磷酸化SlZAT5的Ser-65位點(diǎn),調(diào)控其對(duì)下游成熟相關(guān)基因的轉(zhuǎn)錄抑制活性。
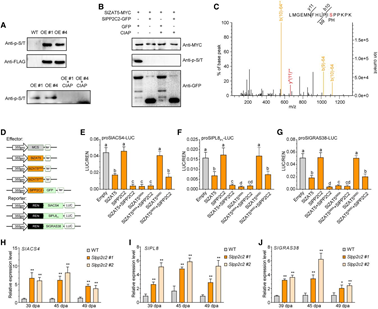
圖5. SlPP2C2對(duì)SlZAT5去磷酸化作用對(duì)SlZAT5轉(zhuǎn)錄活性的影響。

- Plant Physiol:DAP-seq助力揭示番茄果實(shí)成熟調(diào)控新機(jī)制
- DAP-seq助力解析大麥HvbZIP87基因讓小麥抗病又高產(chǎn)的新機(jī)制
- Plant Physiol | DAP-seq助力揭示楊樹根系抗旱新機(jī)制
- Mol Hortic | DAP-seq助力揭示軟棗獼猴桃果實(shí)變色機(jī)制
- 文章分享ChIP-seq:如何解開擬南芥復(fù)制應(yīng)激謎團(tuán)?
- CUT&Tag 技術(shù)原理及應(yīng)用介紹
- 三大技術(shù)如何聯(lián)手揭開PAC1腫瘤免疫的神秘面紗?
- 單細(xì)胞空間轉(zhuǎn)錄組技術(shù)助力發(fā)現(xiàn)腫瘤癌前病變關(guān)鍵細(xì)胞生態(tài)位
- 10x新品發(fā)布網(wǎng)絡(luò)研討會(huì):Visium HD 3’空間分析方案
- Oxford Nanopore明日直播:納米孔測(cè)序大會(huì)亞太專場(chǎng)
- 10x邀您報(bào)名Visium和Xenium數(shù)據(jù)分析研討會(huì)(北京場(chǎng))
- 第三屆中國國際生殖醫(yī)學(xué)健康大會(huì)(北京站)通知
- 10x Genomics技術(shù)方案更新(4月),新手冊(cè)邀您領(lǐng)取
- 10x邀您參加單細(xì)胞與空間多組學(xué)前沿峰會(huì)-北京站
- 真邁FASTASeq 300 Dx獲批NMPA三類醫(yī)療器械注冊(cè)證
- 第五屆多組學(xué)科研與臨床應(yīng)用大會(huì)五一福利大放送


Copyright(C) 1998-2025 生物器材網(wǎng) 電話:021-64166852;13621656896 E-mail:info@bio-equip.com